
כולנו לימדנו כילדים שיש חושים 5: מראה, טעם, צליל, ריח ומגע. ארבעת החושים הראשונים משתמשים באיברים ברורים וברורים, כגון העיניים, בלוטות הטעם, האוזניים והאף, אבל איך בדיוק תחושת הגוף נוגעת בדיוק? מגע הוא מנוסה על כל הגוף, הן מבפנים והן מבחוץ. אין איבר אחד מובחן האחראי על מגע חישה. במקום זאת, יש קולטנים זעירים, או קצות עצבים, סביב הגוף כולו אשר תחושת מגע היכן הוא מתרחש ושולח אותות למוח עם מידע לגבי סוג של מגע שהתרחש. כמו ניצן טעם על הלשון מזהה טעם, mechanoreceptors הם בלוטות בתוך העור ועל איברים אחרים לזהות תחושות של מגע. הם ידועים מכני כי הם נועדו לזהות תחושות מכניות או הבדלים בלחץ.
תוכן
תפקיד מכני
אדם מבין כי הם חוו תחושה ברגע שהאורגן האחראי על גילוי החוש הספציפי הזה שולח מסר למוח, שהוא האיבר העיקרי שמעבד ומארגן את כל המידע. הודעות נשלחות מכל אזורי הגוף למוח באמצעות חוטים המכונה נוירונים. ישנם אלפי נוירונים קטנים אשר מסתעפים לכל חלקי הגוף האנושי, ועל קצות רבים של נוירונים אלה הם mechanoreceptors. כדי להדגים מה קורה כאשר אתה נוגע באובייקט, נשתמש בדוגמה.
דמיינו אדמות יתושים על הזרוע. המתח של החרק הזה, כל כך קל, מגרה את מכני-החושים באזור המסוים הזה של הזרוע. אלה מכניטורים לשלוח הודעה לאורך הנוירון הם מחוברים. הנוירון מתחבר כל הדרך אל המוח, אשר מקבל את המסר שמשהו נוגע לגוף שלך במקום המדויק של mechanoreceptor מסוים ששלח את ההודעה. המוח יפעל עם עצה זו. אולי יגיד לעיניים להסתכל באזור הזרוע שזיהה את החתימה. וכשהעיניים מספרות למוח שיש יתוש על הזרוע, המוח יכול להגיד את היד כדי להעיף אותו במהירות. כך פועלים מכניסטים. מטרת המאמר להלן היא להפגין, כמו גם לדון בפירוט בארגון תפקודי דטרמינציות מולקולריות של mechanoreceptors.
מגע Sense: ארגון פונקציונלי וקביעת מולקולרית של קולטנים רגישים למגע
תַקצִיר
Comporeceptors עורית ממוקמים בשכבות שונות של העור שבו הם מזהים מגוון רחב של גירויים מכניים, כולל מברשת אור, מתיחה, רטט ולחץ מזיק. מגוון זה של גירויים מותאמת על ידי מגוון רחב של mechanoreceptors מיוחדים המגיבים דפורמציה עורית בדרך מסוימת וממירים גירויים אלה מבנים מוחיים גבוהים. מחקרים על פני מכני אנספורקטורים וקצות עצביים סנסוריים חושי גנטית מתחילים לחשוף מנגנוני מגע מגע. עבודה בתחום זה סיפקה לחוקרים הבנה מעמיקה יותר של ארגון המעגל העומד מאחורי תפיסת המגע. ערוצי יון רומן התפתחו כמו מועמדים להולדת מולקולות ומאפיינים של זרמים מגודרים מכנית שיפור ההבנה שלנו של מנגנוני הסתגלות לגירויים מישוש. סקירה זו מדגישה את ההתקדמות שבוצעה אפיון המאפיינים הפונקציונליים של mechanoreceptors בעור שעיר גלאברוס וערוצי יון המזהים תשומות מכניות צורה הסתגלות mechanoreceptor.
מילות מפתח: מכניקה, ערוץ מכני-רגיש, כאב, עור, מערכת סומטו-סנסורית, מגע
מבוא
מגע הוא זיהוי של גירוי מכני המשפיעים על העור, כולל גירויים מכניים מזיקים ומזיקים. זוהי תחושה חיונית להישרדות ולהתפתחות של יונקים ושל בני אדם. מגע של חפצים מוצקים ונוזלים עם העור נותן מידע חיוני למערכת העצבים המרכזית המאפשרת חקר והכרה של הסביבה ויוזמת תנועה או תנועת יד מתוכננת. מגע הוא גם מאוד חשוב עבור החניכות, קשרים חברתיים ומיניות. תחושת המגע היא החוש הפחות פגיע, למרות שזה יכול להיות מעוות (hyperesthesia, hypoesthesia) בתנאים פתולוגיים רבים. 1-3
תגובות מגע כוללות קידוד מדויק מאוד של מידע מכני. קולטנים מכניים עוריים ממוקמים בשכבות העור השונות, שם הם מזהים מגוון רחב של גירויים מכניים, כולל מברשת קלה, מתיחה, רטט, סטיה של שיער ולחץ מזיק. למגוון הגירויים הזה תואם מערך מגוון של מכני קולטנים מיוחדים המגיבים לעיוות עור באופן ספציפי ומעבירים גירויים אלה למבני מוח גבוהים יותר. נוירונים סומטו-סנסוריים של העור מתחלקים לשתי קבוצות: מכני-רצפטורים בעלי סף נמוך (LTMR) המגיבים ללחץ שפיר ומנגנים-קולטני-סף גבוהים (HTMR) המגיבים לגירוי מכני מזיק. גופי תאי LTMR ו- HTMR שוכנים בתוך גרעיני שורש הגבי (DRG) וגרעינים חושי גולגולתי (גרעיני המידה הטריגמינל). סיבי עצב הקשורים ל- LTMR ו- HTMR מסווגים כ- A? -, A? - או C- סיבים בהתבסס על מהירויות ההולכה הפוטנציאליות שלהם. סיבי C אינם ממינליים והם בעלי מהירויות הולכה איטיות ביותר (~ 2 מ 'לשנייה), ואילו A? ו- A? סיבים הם מיאליניזציה קלה ובכבדות, ומציגים מהירויות הולכה בינוניות (~ 12 מ '/ ש') ומהירות (~ 20 מ '/ ש'), בהתאמה. LTMRs מסווגים גם כתגובות איטיות או במהירות (SA- ו- RA-LTMRs) בהתאם לשיעורי ההסתגלות שלהם לגירוי מכני מתמשך. הם נבדלים עוד יותר על ידי איברי הקצה העוריים שהם מעצבנים והגירויים המועדפים עליהם.
היכולת של mechanoreceptors לזהות רמזים מכניים מסתמך על נוכחותם של ערוצי יון mechanotransducer כי להפוך במהירות כוחות מכניים לתוך אותות חשמליים depolarise שדה פתוח. זה depolarisation מקומי, שנקרא פוטנציאל קולטן, יכול לייצר פוטנציאל פעולה כי להתפשט לעבר מערכת העצבים המרכזית. עם זאת, תכונות של מולקולות כי לתווך מכאוטראנצ'וקציה הסתגלות לכוחות מכניים להישאר ברורים.
בסקירה זו אנו מספקים סקירה כללית על תכונות המכונא-קולטנים של יונקים במגע מזיק ומזיק בעור השעיר והלקות. אנו רואים גם את הידע העדכני אודות תכונותיהם של זרמים מגודרים מכנית בניסיון להסביר את מנגנון ההסתגלות של מכני הקולט. לבסוף, אנו סוקרים את ההתקדמות האחרונה שזיהו תעלות יונים וחלבונים נלווים האחראיים לייצור זרמים מכותים.
מגע מזויף
LMRs הקשורים זקיק השיער
זקיקי השיער מייצגים שיער מייצר מיני-איברים המזהים מגע קל. סיבים הקשורים זקיקי השיער להגיב תנועה שיער הכיוון שלה על ידי ירי רכבות של פוטנציאל פעולה עם תחילת והסרה של הגירוי. הם במהירות התאמת קולטנים.
חתול וארנב. במעיל חתולים וארנבות ניתן לחלק את זקיקי השיער בשלושה סוגי זקיקי שיער, שיער הפוך, שיער הגארד והטילוטריץ '. שערות הפוך (שיער תחתון, צמר, וולוס) 4 הן השערות הרבות ביותר, הקצרות והמשובחות ביותר של המעיל. הם גלי, חסרי צבע והגיחו בקבוצות של שתיים עד ארבע שערות מתוך פתח משותף בעור. שערות השומר (מונוטריצ'ים, שמיעה, שיניים) 4 מעוקלות מעט, פיגמנטיות או לא פיגמנטיות, והגיחו באופן יחיד מפי זקיקיהן. הטילוטריצ'ים הם הכי פחות רבים, השערות הארוכות והעבות ביותר .5,6 הם פיגמנטיים או לא פיגמנטיים, לפעמים שניהם והגיחו באופן יחיד מתוך זקיק המוקף בלולאה של כלי דם נימים. סיבי החישה המסופקים לזקיק השיער ממוקמים מתחת לבלוטת החלב ומיוחסים ל- A? או סיבי A? -LTMR .7
בהנחה קרובה לפיר השיער למטה, ממש מתחת לרמה של בלוטת החלב הוא הטבעת של סיומות פיילוט Ruffini משוננת. קצות העצבים הסנסוריים האלה ממוקמים בקורס ספירלי סביב פיר השיער בתוך רקמת החיבור המרכיבה את זקיק השיער. בתוך זקיק השיער, יש גם קצות עצבים חופשיים, חלקם יוצרים מכונאי. לעתים קרובות, לגעת בעורקים (ראה עור glabrous) הם סביב אזור הצוואר של זקיק tilotrich.
מאפייני קצות העצבים המיאליניים בעור שעיר של חתול וארנבון נחקרו באופן אינטנסיבי בתקופת 1930-1970 (סקירה ב- Hamann, 1995) .8 למרבה הפלא, בראון ואיגו, למדו 772 יחידות עם סיבי עצב מואליניים עצבים עצביים של החתול. וארנב, סיווגו תגובות בשלושה סוגי קולטן המתאימים לתנועות שערות למטה (קולטני סוג D), שיער משמר (קולטני סוג G) ושיער טיילוטריץ '(קולטן מסוג T) .9 כל התגובות הסיביות העצביות המובאות נקבצו יחד. בקולטן המותאם במהירות של סוג I (RA I) על ידי התנגדות לקולטן הפאציאני בשם RA II. מכונאי קולטני RA I מגלים מהירות של גירוי מכני ובעלי גבול חד. הם אינם מזהים וריאציות תרמיות. ברג'ס ואח '. תיאר גם קולטן שדה המסתגל במהירות המגיב בצורה אופטימלית לליטוף העור או לתנועה של מספר שערות, אשר יוחס לגירוי של קצות פילו-רופי. אף אחת מתגובות זקיק השיער לא יוחסה לפעילות סיבי C
עכברים. בעור השעיר הגבי של העכברים, שלושה סוגים עיקריים של זקיקי שיער תוארו: זיגזג (סביב 72%), אוול / אאוצ'ן (סביב 23%) ושומר או טיילוטריך (סביב 5%) 11-14 זיגזג ואול / auchenne שיער זקיקי לייצר את רזה פירים שיער קצר יותר והם קשורים עם בלוטת חלב אחת. המשמר או השיער tylotrich הם הארוך ביותר של סוגי זקיקי השיער. הם מאופיינים על ידי נורת שיער גדולה המשויכת שתי בלוטות החלב. המשמר ו awl / שערות auchene מסודרים דפוס איטרטיבי, קבוע ברווח בעוד שערות זיגזג בצפיפות אוכלוסיות אזורי העור סביב שני סוגי זקיקי השיער הגדולים [איור]. 1 (A1, A2 ו A3)].
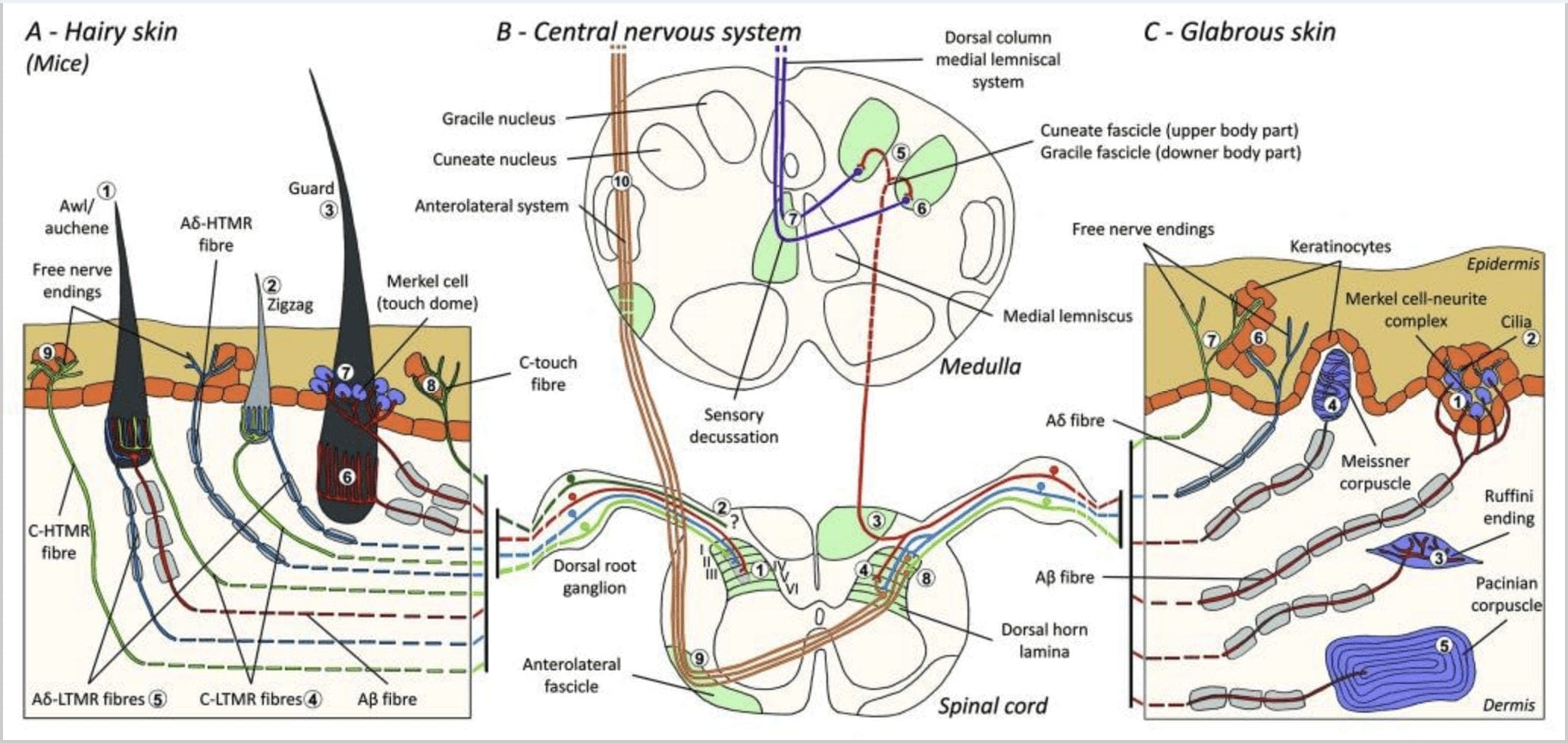
האיור 1. ארגון והקרנות של מכני. בעור שעיר, מברשת קלה ומגע מתגלות בעיקר על ידי העצבנות סביב זקיקי השיער: אובל / אאכן (A1), זיגזג (A2) ושומר (A3). שערות Awl / auchene מעוצבות באופן משולש על ידי קצות אזמל C-LTMR (A4), A? -LTMR ו- A? הסתגלות מהירה- LTMR (A6). זקיקי שיער זיגזג הם פירי השיער הקצרים יותר ומעוצבים על ידי C-LTMR (A4) וגם A? -סופי LTMR ארוכים (A5). זקיקי השיער הארוכים ביותר מונחים על ידי A? הסתגלות מהירה של LTMR אורכי אורך (A6) וקשורות ל- A? התאמה איטית-LTMR של קצות כיפת מגע (A7). התחזיות המרכזיות של כל הסיבים הללו מסתיימות במישורות מובחנות אך חופפות חלקית של קרן הגב של חוט השדרה (C-LTMR ב- lamina II, A? -LTMR ב- lamina III ו- A? -LTMR ב- lamina IV ו- V). התחזיות של LTMR כי innervate זהה או זקיקי השיער הסמוך מיושרים כדי ליצור עמוד צר בעמוד השדרה הגבי השדרה (B1 באפור). רק בעור השעיר, תת-אוכלוסיה של סיבי סיבים חופשיים מסתיימת באפידרמיס ומגיבה למגע נעים (A8). סיבי C-touch אלה אינם מגיבים למגע מזיק ומסע הדרך שלהם עדיין לא ידוע (B2). בעור גס, מגע מזויף מתווך על ידי ארבעה סוגים של LTMRs. תסביך תאי המרקל של מרקל נמצא בשכבת הבסיס של האפידרמיס (C1). מכני קולט זה מורכב מסידור בין תאי מרקל רבים ומסוף עצב מוגדל מ A יחיד? סיבים תזונתיים. תאים Merkel מציג אצבע כמו תהליכים יצירת קשר עם קרטינוציטים (C2). הסוף Ruffini הוא מקומי בדרמיס. זהו קצות חושים מקופלים בצורת סיגר מחוברים ל- A? סיבים (C3). גופת מייזנר מחוברת ל- A? קצה העצבים וממוקם בפפיליות העוריות. זה מכני מוחלטים encorulated מורכב תאים ארוז למטה ארוז מסודרים lamellae אופקי מוקף רקמת חיבור (C4). Corcuscle Pacinian הוא mechanoreceptor עמוק. יחיד A אחד? קצה העצבים הבלתי מיאליני מסתיים במרכז הגופה הגדולה הביצית הזו העשויה ממלות קונצנטריות. התחזיות של סיבי A? -LTMR בחוט השדרה מחולקים לשני ענפים. הסניף המרכזי העיקרי (B3) עולה בחוט השדרה בגופם הקונסטי של הגבינה או בגוש הקשקשים (B5) על רמת medulla, כאשר ה- afferents העיקרי עושה את הסינפסה הראשונה (B6). נוירונים משניים לבצע decusation חושית (B7) כדי ליצור דרכי על lmniscus המדיאלי אשר עולה דרך גזע המוח אל המוח התיכון, במיוחד התלמוס. חטיבה משנית של LTMR מסתיימת בקרן הגבי ב lamina II, IV, V ומפריעה להעברת הכאב (B4). מגע מזיק הוא זוהה על ידי עצב הסוף החופשי האפידרמיס של שניהם שעיר (A9) ו עור glabrous (C7). קולטני מכני אלה הם הסוף של A? -HTMR ו- C-HTMR במגע קרוב עם קרטינוציטים שכנים (C6). A-hTMR מסתיים בלמידה I ו- V; C-HTMR מסתיים בלמידה I ו- II (B8). ברמה של עמוד השדרה הגבי של עמוד השדרה, ה- HTMRs העיקרי של ה- HTMRs יוצר סינפסות עם נוירונים משניים החוצים את קו האמצע ומטפסים אל מבנה המוח הגבוה יותר במסך האנטרוליטרי (B9, B10).
לאחרונה, ג'ינטי ומשתפי הפעולה השתמשו בשילוב של תיוג מולקולרי-גנטי וגישות מעקב רטרוגרדיות סומטוטופיות כדי לדמיין את ארגון הקצוות האקסונליים ההיקפיים והמרכזיים של ה- LTMR בעכברים .15 ממצאיהם תומכים במודל שבו מאפיינים בודדים של גירוי מישוש מורכב הם מופק על ידי שלושת סוגי זקיקי השיער ומועבר באמצעות פעילויות שילובים ייחודיים של סיבי A? -, A? - ו- C אל קרן הגב.
הם הראו כי התיוג הגנטי של נוירונים DRG של טירוזין הידרוקסילאז חיובי (TH +) מאפיין אוכלוסייה של נוירונים חושיים לא קופצניים, בעלי קוטר קטן ומאפשרים הדמיה של קצות היקפיים C-LTMR בעור. באופן מפתיע, הענפים האקסונליים של C-LTMRs בודדים התגלו ויוצרים קצות אורך אורכיים אשר קשורים באופן אינטימי לזגזג (80% מהקצות) ו- awl / auchene (20% מהקצות), אך לא לזקיקי השיער של הטילוטריץ '(איור 1 א). 4 (A15)]. זמן רב נחשב לקצות האורך האורכיים שייכים אך ורק ל- A? -LTMR ולכן לא היה צפוי שסיומות ה- C-LTMR יהוו קצות אורך אורכיים .2 C-LTMR אלה הם בעלי התאמת ביניים בהשוואה להסתגלות האיטית והמהירה. קולטני מכאניים מיאלינים [איור 1 (CXNUMX)].
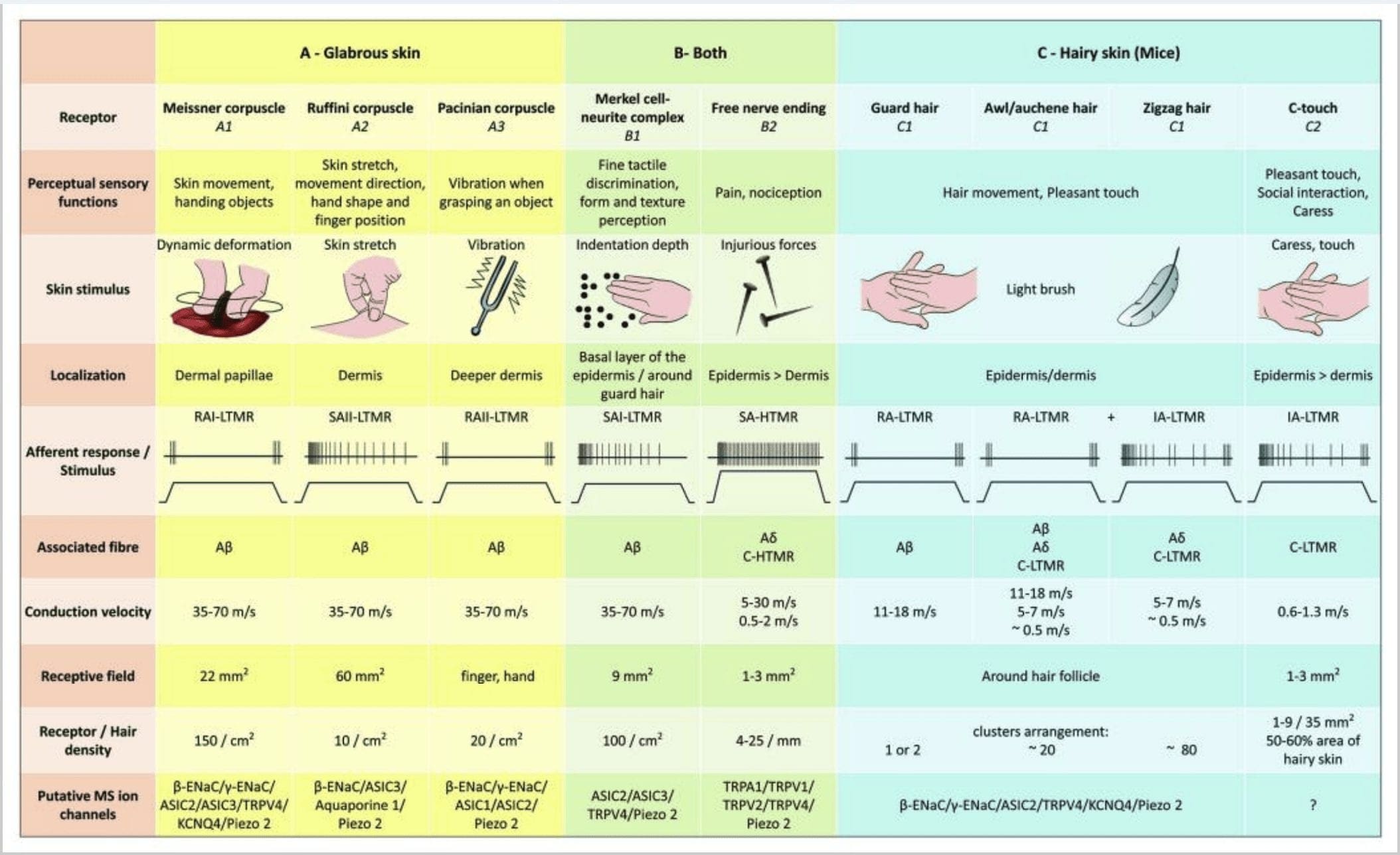
האיור 2. קולטני מישוש אצל יונקים: קולטני מישוש עוריים מתמיינים במגע מזיק הנתמך על ידי קולטנים מרובים עם סף מכני נמוך (LTMR) בעור קרח ושעיר ומגע מזיק הנתמך על ידי קולטני סף מכניים גבוהים (HTMR). הם מרכיבים קצות ללא עצבים שמסתיימים בעיקר באפידרמיס. (א) עור ללא קרח. A1: גופות מייזנר מזהות תנועת עור והחלקת אובייקט ביד. הם חשובים למסירת חפצים וזריזות. קולטנים מסתגלים במהירות לגירוי, מחוברים ל- A? סיבים ודלילים לסיבי C ובעלי שדה קולטן גדול. A2: גופי רופיני מזהים מתיחת עור וחשובים לזיהוי מיקום האצבע וחפץ המסירה. הקולט מסתגל לאט לגירוי ושומר על פעילות כל עוד הגירוי הוחל. קולטנים מחוברים ל- A? סיבים ובעלי שדה קליטה גדול. A3: גופות פציניות נמצאות עמוק יותר בדרמיס ומגלות רטט. קולטנים מחוברים ל- A? סיבים; הם מסתגלים במהירות לגירוי ובעלי השדה הקולט הגדול ביותר. (ב) עור שלם. B1: מתחמי תאי מרקל קיימים גם בעור קרום שיער וגם סביב שיער. הם באים לידי ביטוי בצפיפות ביד וחשובים לתפיסת המרקם ולאפליה הטובה ביותר בין שתי נקודות. הם אחראים על דיוק האצבעות. קולטנים מחוברים ל- A? סיבים; הם מסתגלים אט אט לגירוי ובעלי שדה קליטה קצר. B2: HTMRs מגע מגעילים עם התאמה איטית מאוד לגירוי, כלומר, פעילים כל עוד מוחל הגירוי הנוקיספטיבי. הם נוצרים על ידי הקצה העצבי החופשי של A? וסיבי C הקשורים לקרטינוציטים. (ג) עור שעיר. C1: זקיקי שיער קשורים לסוגי השיער השונים. בעכברים שערות השומר הן הארוכות והביטוי הדליל ביותר, הסמל / אוכנה הם בגודל בינוני והזגזג הוא השיער הקטן ביותר והביטוי ביותר. הם מחוברים ל- A? סיבים אבל גם ל- A? וסיבי C-LTMRs לשיער סלסול / אוקסן וזיזאג. הם מזהים תנועת שיער כולל מגע נעים במהלך ליטוף. הם מסתגלים במהירות או עם קינטיקה בינונית לגירוי. C2: קצות העצבים C-touch תואמים תת-סוג של קצה סיבי C עם סוף חופשי המאופיין בסף מכני נמוך. הם אמורים לקודד לתחושה נעימה הנגרמת על ידי ליטוף. הם מסתגלים בינוני לגירוי ובעלי שדה קליטה קצר. תעלות יונים מכאניות-רגישות (MS) המשוערות המתבטאות בקולטני המישוש השונים מסומנות בהתאם לנתונים ראשוניים ומסכמות את ההשערה הנוכחית בהערכה.
אוכלוסיה מרכזית שנייה שזוהתה נוגעת לסופי A? -LTMR בזקיקי Awl / Auchenne ו- zigzag, בהשוואה לזקיק השיער Down של החתול והארנב. ג'ינטי ומשתפי פעולה הראו כי TrkB מתבטא ברמות גבוהות בתת קבוצה של נוירונים DRG בינוני-קוטר. הקלטות תוך-תאיות באמצעות הכנת העצב העצבית של העור של סיבים שכותרתו גילו כי הן מציגות את התכונות הפיזיולוגיות של סיבים שנחקרו בעבר בחתול וארנב: רגישות מכנית נהדרת (Von Frey <0.07 mN), התאמה מהירה של התגובות לגירויים על-סף, הולכה בינונית מהירויות (5.8 - 0.9 מ '/ ש') וקוצים סומא צרים ולא מוזנים .15 A? -LTMR אלה יוצרים קצות אורך אורכיים הקשורים כמעט לכל זיגזג וזקיק שיער זקיק / auchene של תא המטען [איור. 1 (A5)].
לבסוף הם הראו כי הסופים ההיקפיים של התאמה מהירה של A? LTMR יוצרים קצות אורך אורכיים הקשורים למגן (או טילוטריץ ') ולזקיקי שיער awl / auchene [איור 1 (A6)] .15 בנוסף, שערות המשמר קשורות גם למתחם תאי מרקל ויוצר כיפת מגע המחוברת ל- A? התאמת LTMR לאט [איור 1 (A7)].
לסיכום, כמעט כל זקיקי השיער בזגזג הם עצבניים על ידי קצות L-RTM ו- A? -LTMR קצרים אזמליים; שערות awl / auchene מעוצבות שלוש פעמים על ידי A? הסתגלות מהירה של LTMR, A? -LTMR ו- C-LTMR קצוות אורך; זקיקי שיער השומר מוחצנים על ידי A? הסתגלות מהירה של קצוות אורך רוחביים LTMR ואינטראקציה עם A? התאמה איטית- LTMR של קצות כיפת מגע. לפיכך, כל זקיק שיער בעכבר מקבל שילובים ייחודיים ובלתי משתנים של קצות LTMR המתאימים לאברי קצה מכניים-חושיים מובחנים נוירופיזיולוגית. בהתחשב בסידור החוזר של שלושת סוגי השיער הללו, ג'ינטי ומשתפי הפעולה מציעים שעור שעיר יורכב מחזרה חוזרת של יחידה היקפית המכילה, (1) שערות שמירה אחת או שתיים הממוקמות במרכז, (2) ~ 20 שערות שכבה / עורן סביב ) ~ 3 שערות זיגזג מפוזרות [איור 80 (C2)].
הקרנת חוט השדרה. התחזיות המרכזיות של A? הסתגלות מהירה של LTMRs, A? -LTMRs ו- C-LTMRs מסתיימות בלמידות שונות, אך חופפות חלקית (II, III, IV) של קרן הגב של חוט השדרה. בנוסף, המסופים המרכזיים של LTMRs המעצבנים את אותם זקיקי שיער או סמוכים בתוך יחידת LTMR היקפית מיושרים ליצירת עמוד LTMR צר בקרן הגב של חוט השדרה [איור 1 א). 1 (B2,000)]. לפיכך, נראה כי טריז או טור של קצות חושים ראשוניים מאורגנים סומטוטופית בקרן הגב מייצגים את יישור התחזיות המרכזיות של A? -, A? - ו- C-LTMRs שמפנים את אותה יחידה היקפית ומגלים מכני גירויים הפועלים על אותה קבוצה קטנה של זקיקי שערות. בהתבסס על מספר השערות, השמש / אוצ'ין והזיגזג של תא המטען והגפיים והמספרים של כל תת-סוג LTMR, ג'ינטי ומשתפי פעולה מעריכים כי קרן הגב של העכבר מכילה 4,000 - 15 עמודות LTMR, המקבילות למספר המשוער של היקפי היקפי. יחידות LTMR .XNUMX
יתר על כן, אקסונים של תת-סוגים מסוג LTMR קשורים זה לזה באופן הדוק, עם השלכות משולבות וקצות אזמלים אינטגרטיביים המעצבנים את אותו זקיק השיער. בנוסף, מכיוון ששלושת סוגי זקיקי השיער מציגים צורות, גדלים והרכבים סלולריים שונים, הם עשויים להיות בעלי תכונות כיוונון סטייה או רטט מובהקות. ממצאים אלה עולים בקנה אחד עם מדידות נוירופיזיולוגיות קלאסיות אצל החתול והארנב המצביעות על כך ש A? ניתן להפעיל דיפרנציאלי RA-LTMR ו- A? -LTMR על ידי סטיה של סוגי זקיקי שיער שונים.
לסיכום, מגע בעור השעיר הוא שילוב של: (1) המספרים היחסיים, התפלגות מרחבית ייחודית ותכונות מורפולוגיות וסטלקטואליות שונות של שלושת סוגי זקיקי השיער; (2) שילובים ייחודיים של קצות משנה תת LTMR הקשורים לכל אחד משלושת סוגי זקיקי השיער; ו (3) רגישויות שונות, מהירות הולכה, דפוסי רכבת ספייק ותכונות הסתגלות של ארבעת המעמדות העיקריים של LTRRs הקשורים זקיק השיער, המאפשרים את מערכת השיער מכני חושית כדי לחלץ ולהעביר CNS את השילובים המורכבים של תכונות המגדירים לגעת.
חינם עצבים Endings LTMRs
באופן כללי, קצות חופשיים של סיבי C בעור הם HTMR, אך אוכלוסיית משנה של סיבי C אינה מגיבה למגע מזיק. תת-קבוצה זו של מולישי סיב C (CT) מישושיים מייצגת סוג מובהק של יחידות מכני קבלת פנים לא מיאליניות בעלות סף נמוך הקיימות בעור השעיר אך לא החלקלק של בני אדם ויונקים [איור 1 א). 8 (A18,19)]. 20,21 CT קשורים בדרך כלל לתפיסה של גירוי מישוש נעים במגע הגוף.
נציגי CT מגיבים לכוחות הזחה בטווח של 0.3 2.5 mN ולכן הם רגישים לעיוות העור כמו רבים מה- A? 19 מאפייני ההסתגלות של חסידי CT הם אפוא בינוניים בהשוואה למנגנוריצפטורים המיאלינטיים המסתגלים לאט ומהיר. השדות הקולטים של תאי CT אנושיים הם בצורתם עגולה או אליפסה. השדה מורכב מתשע עד תשעה נקודות תגובה קטנות המפוזרות על שטח של עד 35 מ"מ. 2.22 קולטני ההומולוג של העכבר מאורגנים בתבנית של טלאים לא רציפים המכסים כ- 50 60% מהשטח בעור השעיר [איור 2 א). 2 (C23)] .XNUMX
עדות מחולים ללא affileents מליטין myileinated עולה כי איתות ב סיבי CT להפעיל את הקורטקס המבודד. מאחר שמערכת זו גרועה בקידוד היבטים מפלים של מגע, אך מתאימה היטב לקידוד מגע איטי ועדין, סיבי CT בעור השעיר עשויים להיות חלק ממערכת לעיבוד היבטים נעימים וחשובים מבחינה חברתית של מגע. יש תפקיד בעיכוב כאב והוצע לאחרונה כי דלקת או טראומה עשויה לשנות את התחושה המועברת על ידי סיבים LTMRs סיבים מגע נעים לכאב. 24
איזה מסלול CT-afferents נסיעות עדיין לא ידוע [איור 3]. 1 (B2)], אך תשומות מישוש נמוכות לתאי הקרנה spinothalamic תועדו, 27 ההלוואות אמינות דוחות של גירעונות עדינים, contralateral של זיהוי מגע בחולים בני אדם בעקבות הרס של מסלולים אלה לאחר chordotomy נהלים. 28
LTMRs ב עור Glabrous
מתחמי תא נוירונים של מרקל וכיפת מגע. מרקל (1875) הייתה הראשונה שנתנה תיאור היסטולוגי של אשכולות של תאי האפידרמיס עם גרעינים גדולים המכונסים, ויצרו קשר עם סיבי עצב מראים. הוא הניח שהם שקעו את תחושת המגע בכך שכינו אותם טסטזלן (תאי מישוש). בבני אדם, מתחמי תאי הנויריט של מרקל מועשרים באזורים רגישים למגע בעור, והם נמצאים בשכבה הבסיסית של האפידרמיס באצבעות, בשפתיים ובאיברי המין. הם קיימים גם בעור שעיר בצפיפות נמוכה יותר. מתחם תא מרקל מורכב מתא מרקל במיקום צמוד לטרמינל עצב מוגדל מ- A מיאלין יחיד? סיבים [איור 1 (C1)] (סקירה בהלטה ומשתפי פעולה) .29 בצד האפידרמיס תאי מרקל מציגים תהליכים דמויי אצבעות המשתרעים בין קרטינוציטים שכנים [איור 1 א). 2 (C30,31)]. תאי מרקל הם תאי אפידרמיס שמקורם בקרטינוציטים. 150 מונח כיפת המגע הונהג בכינוי שמו של הריכוז הגדול של מתחמי תאי מרקל בעור השעיר של כף קדמת החתול. כיפת מגע יכולה להכיל עד 32 תאי מרקל המופנחים על ידי סיב A יחיד? ובבני אדם מלבד סיבי A?, A? וסיבי C היו גם הם באופן קבוע .34-XNUMX
גירוי של קומפלקסים של תאי מרק נוירוט מביא לתגובות סוג I (SA I) המתאימות לאט, שמקורן בשדות קולטני ניקוד עם גבולות חדים. אין פריקה ספונטנית. מתחמים אלה מגיבים לעומק הכניסה של העור ויש להם את הרזולוציה המרחבית הגבוהה ביותר (0.5 מ"מ) של מכני הקולטנים העוריים. הם משדרים תמונה מרחבית מדויקת של גירויי מישוש ומוצעים להיות אחראים לאפליה של צורה ומרקם. 2 (B1)]. עכברים נטולי תאי מרקל אינם יכולים לזהות משטחי מרקם בכפות רגליהם בזמן שהם עושים זאת באמצעות שפם
בין אם תא מרקל, הנוירון החושי או שניהם הם אתרי תמרוק-מכני הוא עדיין עניין של ויכוח. בחולדות, הרס פוטוטוקסי של תאי מרקל מבטל את תגובת ה- SA I.36 בעכברים עם תאים מדוכאים גנטית, תגובת ה- SA I שנרשמה בהכנת ex / vivo עור / עצב נעלמה לחלוטין, והדגימה כי תאי מרקל נדרשים לקידוד נכון של מרקל. תגובות קולטן .37 עם זאת, גירוי מכני של תאי מרקל מבודדים בתרבית על ידי לחץ מונעי אינו מייצר זרמים מגודרים מכנית. 38,39 קרטינוציטים עשויים למלא תפקיד חשוב בתפקודו הרגיל של מתחם תא המירוי. התהליכים דמויי האצבעות של מרקל יכולים לנוע עם דפורמציה של העור ותנועת תאי האפידרמיס, וזה עשוי להיות הצעד הראשון של התמרה מכנית. ברור שהתנאים הנדרשים לחקר רגישות מכאנו של תאי מרקל טרם נקבעו.
קצות רופיני. קצות רופיני הם קצות חושים מקופלים בצורת סיגר דקים המחוברים ל- A? קצות עצבים. קצות רופיני הם גלילי רקמת חיבור קטנים המסודרים לאורך גדילי קולגן עורי המסופקים על ידי סיב עצבי מיאלין אחד עד שלושה ממדי 4 מטר. עד שלושה גלילים בעלי כיוון שונה בדרמיס עשויים להתמזג ליצירת קולטן אחד [איור 6 א). 1 (C3)]. מבחינה מבנית, קצות רופיני דומים לאברי גיד של גולגי. הם באים לידי ביטוי באופן כללי בדרמיס וזוהו כמכני קולטניים עוריים מסוג II (SA II) המתאימים לאט. על רקע פעילות עצבית ספונטנית, פריקה סדירה המסתגלת לאט מתעוררת על ידי גירוי מכני נמוך מאונך הנשמר או בצורה יעילה יותר על ידי מתיחה בעור. תגובת SA II מקורם בשדות קליטה גדולים עם גבולות לא ברורים. קולטני רופיני תורמים לתפיסת כיוון תנועת האובייקט דרך דפוס מתיחת העור [איור 2 א). 2 (AXNUMX)].
בעכברים, SA אני ו II תגובות יכול להיות מופרד electrophysiologically ב vivo לשעבר עור עצבני הכנה. 40 Nandasena ומשתפי פעולה דיווחו immunolocalization של אקואפורן 1 (AQP1) ב endontal Ruffini סופים של חולדות העכברים המציע AQP1 מעורב ב תחזוקת האיזון האוסמוטי הדנטאלי הנחוץ עבור המוטרו-טרנספורמציה. 41 סיומות הקצינים של Rudini גם הביעו את הערוץ היוני מכני החיסוני ASIC3.42
שרירי מייסנר. גופי גוף מייזנר ממוקמים בפפיליות העור של הזגגות, בעיקר בכפות הידיים ובכפות הרגליים, אך גם בשפתיים, בלשון, בפנים, בפטמות ובאיברי המין. מבחינה אנטומית, הם מורכבים מקצה עצבי מקופל, כשהקפסולה עשויה תאים תומכים שטוחים המסודרים כממלים אופקיים המוטבעים ברקמת החיבור. יש סיב עצבי אחד A? מציאים מחוברים לכל גופה [איור 1 (C4)]. כל דפורמציה גופנית של הגופנית מפעילה מטח של פוטנציאל פעולה המפסיק במהירות, כלומר, הם מתאימים קולטנים במהירות. כאשר מסלקים את הגירוי, הגופה מחזירה את צורתה ותוך כדי כך מייצרת מטח נוסף של פוטנציאל פעולה. בשל מיקומם השטחי בדרמיס, גופות אלה מגיבות באופן סלקטיבי לתנועת העור, לזיהוי מישוש של החלקה ורטט (20 הרץ). הם רגישים לעור דינמי - למשל, בין העור לאובייקט שמטופל בו [איור 40 א). 2 (A1)].
גופים פאציניים. גופי גוף פאציניאניים הם קולטני המנגנון העמוקים יותר של העור והם המכני קולטני העור המעוגן הרגיש ביותר בתנועת העור. גופי גוף גדולים אלה (אורך 1 מ"מ) עשויים מממלות קונצנטריות של רקמת חיבור סיבית ופיברובלסטים מרופדים על ידי תאי שוואן שטוחים ומשתנים באים לידי ביטוי בדרמיס העמוק .43 במרכז הגופה, בחלל מלא נוזלים הנקרא נורה פנימית. , מסיים A יחיד אחד? קצה עצבי לא מיאליני מציא [איור 1 (C5)]. יש להם שדה קולט גדול על פני העור עם מרכז רגיש במיוחד. ההתפתחות והתפקוד של כמה סוגי מכני קולטנים מסתגלים במהירות מופרעים בעכברים מוטנטים c-Maf. בפרט, גופות פאציניאניות ניוון מאוד
גופיות פאציניאניות מציגות הסתגלות מהירה מאוד בתגובה לכניסת העור, הפרשת העצבים II (RA II) המסתגלת במהירות המסוגלות לעקוב אחר תדירות גבוהה של גירויים רטטיים, ומאפשרות תפיסה של אירועים רחוקים באמצעות תנודות מועברות .45 מציאים מגיבים לכניעה מתמשכת עם פעילות חולפת בתחילת וקיזוז הגירוי. הם נקראים גם גלאי תאוצה מכיוון שהם יכולים לזהות שינויים בחוזק הגירוי, ואם קצב השינוי בגירוי משתנה (כפי שקורה בתנודות), תגובתם הופכת להיות פרופורציונאלית לשינוי זה. גופות פאציניאניות חשות שינויי לחץ גסים ובעיקר רעידות (150 300 הרץ), שאותן הם יכולים לזהות אפילו סנטימטרים משם [איור. 2 (A3)].
תגובה טונית נצפתה ב corpuscle Pacinian decapsulated.46 בנוסף, שלמות Pacinian שלם להגיב עם פעילות מתמשכת במהלך גירויים הזחה קבועים, ללא שינוי סף מכני או תדר התגובה כאשר איתות GABA בתיווך נחסם בין גליה lamellate וסיום עצב. 47 לכן, את רכיבים שאינם עצביים של corpuscle Pacinian עשויים להיות תפקידים כפולים סינון גירוי מכני כמו גם ב modulated את מאפייני התגובה של נוירון חושי.
תחזיות חוט השדרה. תחזיות ה- A? -LTMR בחוט השדרה מחולקות לשני ענפים. הענף המרכזי העיקרי עולה בחוט השדרה בעמודות הגב האפסילטרליות עד לרמה הצווארית [איור 1 א). 3 (B1)]. ענפים משניים מסתיימים בקרן הגב ברבידות הרביעיות ומפריעים להעברת הכאב, למשל. זה עשוי להחליש כאב כחלק מבקרת השער [איור 4 (B48)] XNUMX
ברמות צוואר הרחם, האקסונים של הסניף הראשי נפרדים בשני חלקים: מערכת קו האמצע כוללת את הקשקש הגרעיני המשדר מידע מהחלק התחתון של הגוף (רגליים ותא המטען), והמערכת החיצונית כוללת את המידע המשולב, של הגוף (זרועות ותא המטען) [איור 3]. 1 (B5)].
מספרי מישוש ראשוניים מבצעים את הסינפסה הראשונה שלהם עם נוירונים מסדר שני במדולה, שם מסתננים סיבים מכל דרכי גרעין באותו שם: הסינפסה של האקסונים החינניים של גרסיל בגרעין החינני והסינפסה של האקסונים של גינון בגרעין ה- cuneate [איור 1 א). 6 (B1)]. נוירונים המקבלים את הסינפסה מספקים את המבקרים המשניים וחוצים את קו האמצע באופן מיידי ויוצרים צינור בצד הנגדי של גזע המוח, הלמניסקוס המדיאלי, שעולה דרך גזע המוח לתחנת הממסר הבאה במוח התיכון, במיוחד בתלמוס [איור . 7 (BXNUMX)].
מפרט מולקולרי של LTMRs. לאחרונה הובהר בחלקו מנגנונים מולקולריים השולטים על פיזור מוקדם של LTMRs. בוראן ומשתפי פעולה הראו כי האוכלוסיות העצביות המבטאות את קולטן ה- Ret טירוזין קינאז (Ret) ואת הקולטן המשותף שלו GFR? 2 בעכברים עובריים E11 13 DRG מבטאות באופן סלקטיבי את גורם השעתוק Mafa.49,50 מחברים אלה מדגימים כי Mafa / נוירונים של Ret / GFR? 2 שנועדו להפוך לשלושה סוגים ספציפיים של LTRM בלידה: נוירונים SA1 המפנים מתחמי תאי מרקל, הנוירונים המסתגלים במהירות המעורבים את גופי מייזנר והסמים המתאימים במהירות (RA I) ויוצרים קצות אמצעיים סביב זקיקי השיער. ג'ינטי ומשתפי פעולה מדווחים גם כי נוירונים של DRG המבטאים את ה- Ret-Early מתאימים במהירות מכני קולטנים מגופי גוף מייזנר, גופי פאציניאן וקצות אמצעיים סביב זקיקי השיער .51 הם עורבים אזורי יעד נפרדים בגרעינים החינניים והקונטיים, וחושפים דפוס ספציפי למודל של מכני-חושי. השלכות אקסונליות עצביות בגזע המוח.
חקירה של מכונאי העור האנושי. טכניקת המיקרונורוגרפיה שתוארה על ידי הגברת וואלבו בשנת 1968 יושמה על מנת לחקור את התנהגות הפריקה של קצות מכאניים רגישים אנושיים המספקים שרירים, מפרקים ועור (ראה סקירה Macefield, 2005) .52,53 רוב המיקרונורוגרפיה של העור האנושית. מחקרים אפיינו את הפיזיולוגיה של מצידי מישוש בעור הזרוח של היד. הקלטות מיקרואלקטרודות מהעצם החציוני והאולנרי אצל נבדקים אנושיים חשפו תחושת מגע שנוצרו על ידי ארבע הכיתות של ה- LTMR: מסיימי מייזנר רגישים במיוחד לליטוף אור על פני העור, ומגיבים לכוחות גזירה מקומיים והחלקות מתחילות או גלויות בשדה הקולט. חסידי פאציניאן רגישים להפליא לחולפים מכניים מהירים. הסובלים מגיבים נמרצות לנשוף מעל שדה הקליטה. גופת פאציניאנית הממוקמת בספרה תגיב בדרך כלל להקשה על השולחן התומך בזרוע. לניסיונות מרקל באופן אופייני יש רגישות דינמית גבוהה לגירויי כניסה המיושמים על אזור נפרד ולעתים קרובות מגיבים בהפרשות במהלך השחרור. אף על פי שאמני רופיני מגיבים לכוחות המופעלים באופן רגיל על העור, מאפיין ייחודי של מצידי SA II הוא יכולתם להגיב גם למתיחות עור רוחביות. לבסוף, ביחידות שיער באמה יש שדות קליטה גדולים או לא סדירים המורכבים מכמה כתמים רגישים המתאימים לשערות בודדות (כל אספקה מציגה ~ 20 שערות).
רגישות מכנית של Keratinocytes
כל גירוי מכני על העור חייב להיות מועבר דרך קרטינוציטים המהווים את האפידרמיס. תאים אלה בכל מקום עשויים לבצע פונקציות איתות בנוסף לתפקידים תומכים או מגן שלהם. לדוגמה, קרטינוציטים מפרישים ATP, מולקולה חשובה איתות חושית, בתגובה לגירויים מכניים ואוסמוטיים. 54,55 שחרור ATP גורם גידול סידן תאיים על ידי גירוי אוטוקריני של קולטנים purinergic. 55 יתר על כן, יש ראיות כי hypotonicity מפעילה את Rho- קינאז מסלול איתות ואת שלאחר מכן F- אקטין מתח סיבים היווצרות המציע כי דפורמציה מכנית של keratinocytes עשוי באופן מכני להפריע לתאים שכן כגון תאים Merkel מגע לא מזיק וסיבים C- סיבים חינם מגע רעיל [איור]. 1 (C6)] 56,57
מגע רעיל
קולטני מכני סף גבוהים (HTMR) הם אפידרמיס C ו- A? קצות עצבים חופשיים. הם אינם קשורים למבנים מיוחדים ונצפים בשני העור השעיר [איור 1 א). 9 (A1)] ועור ללא קרח [איור. 7 (C6)]. עם זאת, המונח של סוף עצבים חופשי צריך להיחשב בזהירות מכיוון שקצות העצבים תמיד נמצאים במיקום צמוד עם תא קרטינוציטים או לנגרנס או מלנוציטים. ניתוח אולטרה-מבני של קצות עצבים מגלה נוכחות של רשתית אנדופלזמית גסה, מיטוכונדריה בשפע ושלפוחית צפופה. קרומים סמוכים של תאי האפידרמיס מעובים ודומים לקרום פוסט סינפטי ברקמות העצבים. שים לב כי האינטראקציות בין קצות העצבים לתאי האפידרמיס עשויות להיות דו כיווניות מאחר ותאי האפידרמיס עשויים לשחרר מתווכים כ- ATP, אינטרלוקין (IL10, IL2) וברדיקינין ולהפך קצות עצבים פפטידרגיים עשויים לשחרר פפטידים כגון CGRP או חומר P הפועל על תאי האפידרמיס. HTMRs מכילים nociceptors mechano נרגשים רק על ידי גירויים מכניים מזיקים ו nociceptors polymodal המגיבים גם לחום מזיק וכימיקל אקסוגני [איור. 2 (B58)] XNUMX
סיבים הולמים של HTMR מסתיימים על עצבי הקרנה בקרן הגב של חוט השדרה. A-HTMRs יוצרים קשר עם נוירונים מהסדר השני בעיקר ב lamina I ו- V, ואילו C-HTMRs מסתיימים ב lamina II [איור. 1 (B8)]. נוירונים nociceptive מסדר שני מוקרנים לצד הבקרה של חוט השדרה ועולים בחומר הלבן ויוצרים את המערכת האנטר-צדדית. נוירונים אלו מסתיימים בעיקר בתלמוס [איור 1 (B9 ו- B10)].
מכנו-זרמים של נוירונים סומטו-סנסוריים
המנגנונים של הסתגלות איטית או מהירה של מכאנספטורים עדיין לא הובהרו. לא ברור באיזו מידה הסתגלות מכאנספקטור מסופקת על ידי הסביבה הסלולרית של הסוף העצבי הסנסורי, התכונות המהותיות של הערוצים הנשלטים מכנית ומאפייני ערוצי יון המצוידים במיקוד אקסונלי בנוירונים סנסוריים (איור 2). עם זאת, ההתקדמות האחרונה באפיון של זרמים מגודרים מכנית הוכיחה כי סוגים שונים של ערוצים מכניים נמצאים במעבדי DRG ויכולים להסביר היבטים מסוימים של הסתגלות של מכני.
הקלטה חוץ גופית במכרסמים הראתה כי הסומה של נוירונים DRG הוא באופן מהותי רגיש מכני ומביע ביטוי זרמי מכאנו מכותיים. 59-64 גדוליניום ואדום רותניום חוסמים לחלוטין זרמים מכאניים-רגישים, ואילו סידן ומגנזיום חיצוניים, בריכוזים פיזיולוגיים, כמו גם עמילוריד ובנזמיל, גורמים לחסימה חלקית. 60,62,63 FM1-43 משמש כחסם מתמשך, והזרקת FM1-43 לכף האחורית של העכברים מפחיתה את הרגישות לכאב במבחן רנדל סליטו ומגדילה את סף נסיגת הכף המוערך. עם שערות פון פריי
בתגובה לגירוי מכני מתמשך, זרמים מכניים רגישים יורדים באמצעות סגירה. בהתבסס על קבועי הזמן של הריקבון הנוכחי, הבחינו בארבעה סוגים נפרדים של זרמים רגישים-מכניים: התאמת זרמים מהירה (~ 3 אלפיות שנייה), התאמת זרמים בינוניים (~ 6 אלפיות שנייה), התאמת זרמים איטית (~ 15 עד 30 אלפיות שנייה) ) וזרמים מסתגלים לאט לאט (~ 200 אלפיות שניים) .300 כל הזרמים הללו קיימים עם שכיחות משתנה בנוירונים DRG חולדות המעצבנים את העור הזפה של כף האחורית .1000
ניתן לקבוע את הרגישות המכנית של זרמים רגישים מכאניים על ידי הפעלת סדרת גירויים מכניים מצטברים, המאפשרת ניתוח גירוי-זרם מפורט יחסית. 66 מערכת היחסים בין הגירוי הנוכחי היא בדרך כלל סיגמואידית, והמשרעת המרבית של הזרם נקבעת על ידי מספר ערוצים שפתוחים בו זמנית .64,67 מעניין לציין כי הדיווח על הזרם המכני-רגיש המהיר מציג נקודת אמצע מכנית נמוכה וחצי הפעלה בהשוואה לזרם המכני-רגיש במיוחד המתאים לאט. 63,65
נוירונים חושיים עם פנוטיפים שאינם נוסיצפטיבים מעדיפים להתאימם במהירות לזרמים מכאוסוסיטיביים עם סף מכני נמוך. 60,61,63,64,68 לעומת זאת, זרמים מכוסוסינסטיבים איטיים ומתאימים לאיטי מדווחים לעתים על תאים לא-נוציפטיבים. סף מכני שונים לראות LTMRs ו HTMRs in vivo. למרות אלו ניסויים במבחנה צריך להילקח בזהירות, תמיכה בנוכחות של soma של נוירונים DRG של מכני נמוך ו-סף מכני גבוה ניתנה גם על ידי גירוי מבוססי למתוח רדיאלי של נוירונים חושי עכבר תרבותי. 64,68 פרדיגמה זו חשפה שני האוכלוסיות העיקריות של נוירונים רגישים למתוח, אחד המגיב משרעת גירוי נמוך ועוד אחד סלקטיבי מגיב משרעת גירוי גבוהה.
לתוצאות אלו יש השלכות מכניסטיות, אך ספקולטיביות: הסף המכני של הנוירונים החושתיים עשוי להיות קשור לארגון הסלולר של המנגנון, אך הוא עשוי לשכב במאפייני ערוצי היונים הנשלטים מכנית.
המנגנונים העומדים בבסיס דה-סנסיטיזציה של זרמי קטיון מכניים רגישים בתאי עצב DRG עכברושים נפרמו לאחרונה .64,67 זה נובע משני מנגנונים מקבילים המשפיעים על מאפייני הערוץ: הסתגלות והפעלה. הסתגלות דווחה לראשונה במחקרי תאי שיער שמיעתיים. ניתן לתאר זאת באופן מבצעי כתרגום פשוט של עקומת ההפעלה של ערוץ המתמר לאורך ציר הגירוי המכני .70-72 הסתגלות מאפשרת לקולטנים חושיים לשמור על רגישותם לגירויים חדשים בנוכחות גירוי קיים. עם זאת, לא ניתן להפעיל מחדש חלק ניכר מהזרמים המכאניים-רגישים בתאי עצב DRG בעקבות גירוי מכני של מיזוג, מה שמעיד על הפעלת ערוצי מתמר מסוימים .64,67 לכן, אי-הפעלה והתאמה פועלים בד בבד לוויסות זרמים מכאניים-רגישים. שני מנגנונים אלה משותפים לכל הזרמים הרגישים-מכניים המזוהים בתאי עצב DRG בחולדה, מה שמרמז על כך שאלמנטים פיזיקוכימיים קשורים קובעים את קינטיקה של ערוצים אלה .64
לסיכום, קביעת המאפיינים של זרמים מכניים-רגישים אנדוגניים במבחנה היא מכרעת במסע לזהות מנגנוני התמרה ברמה המולקולרית. השונות שנצפתה בסף המכני ובקינטיקה המותאמת של הזרמים השונים המכאניים בנוירונים של DRG מצביעים על כך שתכונות פנימיות של תעלות יונים עשויות להסביר, לפחות באופן חלקי, קינטיקה של סף מכני והתאמה של מכני הקולטנים שתוארו בעשורים 1960. 80 באמצעות תכשירים ex vivo.
חלבונים מכני
זרמי יון מכאנסנסטיביים בנוירונים סומטו-סנסוריים מאופיינים היטב, לעומת זאת, מעט ידוע על זהותן של מולקולות המווכות את מכנו-טרנספורמציה ביונקים. מסכים גנטיים ב תסיסנית ו C. elegans זיהו מולקולות המועמד mechanotransduction המועמד, כולל TRP ו degenerin / epithelial Na + ערוץ (Deg / ENaC) משפחות. 73 ניסיונות אחרונים להבהיר את הבסיס המולקולרי של mechanotransduction ביונקים התמקדו במידה רבה homologs של המועמדים האלה . בנוסף, רבים מהמועמדים האלה נמצאים במיקרוספטקטורים עוריים ונוירונים סומטוסנסוריים (איור 2).
חומצה חישה יון ערוצים
ASICs שייכים לתת-קבוצה מחוברת פרוטונים ממשפחת ערוצי ה- Na + degenerin אפיתל .74 שלושה מבני משפחת ASIC (ASIC1, ASIC2 ו- ASIC3) באים לידי ביטוי במנגנון-קולטנים ובנוסצפטורים. תפקידם של ערוצי ASIC נחקר במחקרי התנהגות באמצעות עכברים עם מחיקה ממוקדת של גני ערוץ ASIC. מחיקת ASIC1 אינה משנה את תפקודם של מכני קולטניים עוריים אלא מגבירה את הרגישות המכנית של מביאים המעורבים את המעיים .75 עכברי נוקאאוט של ASIC2 מראים רגישות מופחתת של LTMRs עוריים מסתגלים במהירות .76 עם זאת, מחקרים שלאחר מכן דיווחו על היעדר השפעות של דפיקת ASIC2 הן מכאנו-נוסיספציה קרביים והן מכאנו-סנסציה עורית .77 הפרעה ב- ASIC3 מפחיתה את הרגישות של מכאנו של גורמים קרביים ומפחיתה תגובות של HTMRs עוריות לגירויים מזיקים.
ערוץ קולטן חולף
משפחת ה- TRP מחולקת לשישה תת-יונקים ביונקים. 78 כמעט לכל תת-התינוקות של ה- TRP יש חברים המקושרים ל- mechanosensation במגוון מערכות תאים. 79 ב נוירונים חושיים יונקים, לעומת זאת, ערוצי TRP ידועים בעיקר בחשיפת מידע תרמי ודלקת נוירוגנית מתווכת, ורק שני ערוצי TRP, TRPV4 ו TRPA1, היו מעורבים בתגובה המגע. לשבש ביטוי TRPV4 בעכברים יש רק השפעות צנועות על סף mechanosensory אקוטי, אך מפחית באופן משמעותי את הרגישות לגירויים מכניים מזיקים. 80,81 TRPV4 הוא גורם מכריע בעיצוב התגובה של נויציקטיביים נוירונים ללחץ אוסמוטי כדי hyperalgesia מכני במהלך דלקת. 82,83 TRPA1 נראה יש תפקיד hyperalgesia מכני. עכברים חסרים TRPA1 התערוכה כאב רגישות יתר. TRPA1 תורמת התמרה של גירויים מכניים, קרים וכימיים נויצפטורים עצביים סנסוריים אבל נראה כי הוא לא חיוני עבור התמרה שיער תא. 84,85
אין ראיות ברורות המעידות על כך שערוצי TRP וערוצי ASIC המבוטאים ביונקים נשלטים באופן מכני. אף אחד מן הערוצים הללו לא ביטא בצורה חכמה את החתימה החשמלית של זרמים מכנו-סנסוריים שנצפו בסביבתם הטבעית. אין בכך כדי לשלול את האפשרות שערוצי ASICs ו- TRPs הם מכונאי-זרם, בהתחשב באי-ודאות אם ערוץ מכנו-טרנספורמציות עשוי לפעול מחוץ להקשר הסלולרי שלו (ראה סעיף על SLP3).
Piezo חלבונים
Piezo protiens זוהו לאחרונה כמועמדים מבטיחים לחלבונים מכוסמים על ידי Coste ומשתפי פעולה. 86,87 חוליות יש שני חברים Piezo, Piezo 1 ו Piezo 2, הידועה בעבר בשם FAM38A ו FAM38B, בהתאמה, אשר נשמרים היטב לאורך eukaryotes הסלולר מרובות. Piezo 2 הוא שופע ב DRGs, ואילו Piezo 1 הוא בקושי לזיהוי. Piezo- המושרה זרמים מכניים מונעים מעכבות על ידי גדוליניום, רותניום אדום ו GsMTx4 (רעלן מן הטראנטולה Grammostola spatulata). 88 הביטוי של Piezo 1 או Piezo 2 במערכות heterologous מייצרת זרמים mechanosensitive, הקינטיקה של inactivation של Piezo 2 הנוכחי להיות מהר מאשר Piezo 1. בדומה לזרמים האנדוגניים הרגישים, לזרמים תלויי Piezo יש פוטנציאל היפוך סביב 0 mV והם קטיון לא סלקטיבי, עם Na +, K +, Ca2 + ו- Mg2 + כולם חודרים לערוץ הבסיסי. כמו כן, זרמים תלויי פיזו מוסדרים על ידי פוטנציאל ממברנה, עם האטה ניכרת של הקינטיקה הנוכחית על הפוטנציאלים depolarized. 86
חלבונים Piezo הם ללא ספק mechanosensing חלבונים ולשתף תכונות רבות של התאמת במהירות זרמים מכאוסנסטיבי ב נוירונים חושית. טיפול של נוירונים מתורבתים DRG עם Piezo 2 הפרעה קצרה RNA הפחית את חלקם של נוירונים עם התאמת במהירות הנוכחי והקטין את אחוז של נוירונים מכני. נינוקס Transmembrane תחומים ממוקמים בכל חלבוני piezo אבל לא ברור הנקבוביות המכילות מוטיבים או יון חתימות ערוץ כבר מזוהה. עם זאת, עכבר Piezo 86 חלבון מטוהרים מחדש לתוך bilayers השומן האסימטרי ו liposome צורות יון ערוצי רגיש אדום רותניום. 1 צעד חיוני אימות mechanotransduction באמצעות ערוצי Piezo היא להשתמש בגישות vivo כדי לקבוע את החשיבות הפונקציונלית איתות איתות. המידע נמסר בתסיסנית שבה מחיקתו של חבר פיצו אחד הפחיתה את התגובה המכנית לגירויים מזיקים, מבלי להשפיע על מגע נורמלי. 87 למרות שמבנהם נותר להיקבע, משפחה חדשה זו של חלבונים מכני-סנסטיביים היא נושא מבטיח למחקר עתידי, מעבר לגבול של תחושת מגע. לדוגמה, מחקר שנערך לאחרונה על חולים עם אנמיה (xerocytosis תורשתי) מראה את התפקיד של Piezo 89 בשמירה על ההומיאוסטזה נפח כדורית הדם. 1
ערוץ Transmembrane כמו (TMC)
מחקר שנערך לאחרונה עולה כי שני חלבונים, TMC1 ו TMC2, נחוצים עבור mechanotransduction תאים תא. 91 חירשות תורשתית עקב מוטציה TMC1 הגן דווח בעכברים אנושיים ועכברים. 92,93 נוכחות של ערוצים אלה עדיין לא הוכח במערכת סומטו-סנסורי, אבל זה נראה טוב להוביל לחקור.
חלבון סטומאטין כמו 3 (SLP3)
בנוסף ערוצי התמרה, כמה חלבונים אביזר המקושרים לערוץ הוכחו לשחק תפקיד חושי מגע. SLP3 מתבטא נוירונים DRG יונקים. מחקרים באמצעות עכברים מוטציה חסר SLP3 הראו שינוי mechanosensation ו זרמים mechanosentive.94,95 SLP3 פונקציה מדויקת נשאר ידוע. זה עשוי להיות מקשר בין הערוץ מכני רגישות לבין microtubules הבסיסית, כפי שהוצע עבור ג שלה elegans homolog MEC2.96 לאחרונה GR. מעבדה Lewin הציע כי הקושר הוא מסונתז על ידי נוירונים חושי DRG וקישורים ערוץ יון mechanosensitive אל המטריצה תאיים. 97 שיבוש הקישור מבטל הנוכחי RA-mechanosensitive מציע כי כמה ערוצי יון הם מכנית רק כאשר קשורה. זרמי RA-mechanosensitive גם מעוכבים על ידי laminin-332, חלבון מטריקס המיוצר על ידי קרטינוציטים, מחזק את ההשערה של אפנון של זרם מכני של תאים על ידי חלבונים תאיים. 98
K + ערוץ משנה
במקביל לזרמים מכאניים-מינוריים של קטיון, נוכחותם של זרמי K + מכנים מחדש את הממברנה נמצאת תחת חקירה. K + ערוצי בתאי mechanosensitive יכול צעד במאזן הנוכחי ולתרום להגדיר את סף מכני ואת משך הזמן של הסתגלות של mechanoreceptors.
חברי KCNK שייכים לתחום שני הנקבוביות K + ערוץ (K2P) family.99,100 K2P להציג מגוון מדהים של רגולציה על ידי סוכנים סלולריים, פיסיקלית פרמקולוגית, כולל שינויים pH, חום, מתיחה דפורמציה הממברנה. אלה הם K2P פעיל במנוחה פוטנציאל הממברנה. כמה יחידות משנה של KCNK מתבטאות בנוירונים סומטוסנסוריים. 101 KCNK2 (TREK-1), KCNK4 (TRAAK) וערוץ TREK-2 הם בין הערוצים הבודדים שעבורם גייט מכני ישיר על ידי מתיחת קרום הוצג. 102,103
עכברים עם גן KCNK2 משובש הראו רגישות משופרת לחום ולגירויים מכניים קלים, אך סף נסיגה רגיל ללחץ מכני מזיק המופעל על כף הרגל באמצעות מבחן רנדל. סליטו. 104 עכברים חסרי KCNK2 מציגים גם היפרלזיה תרמית ומכנית מוגברת בדלקתיים תנאים. עכברי נוקאאוט של KCNK4 היו רגישים יתר לגירוי מכני קל, ורגישות יתר זו הוגברה על ידי השבתה נוספת של KCNK2.105 רגישות מכאנית מוגברת של עכברי נוקאאוט אלה יכולה לגרום למתיחה להפעיל בדרך כלל גם זרמים מכאניים-רגישים דפולריזמים וגם מקוטבים באופן מתואם, בדומה לחוסר האיזון של זרמים מחוברי מתח דפולריזמים וממולבים מחדש.
KCNK18 (TRESK) הוא תורם מרכזי למוליכות K + המווסתת את פוטנציאל הממברנה המנוחה של נוירונים סומטו-סנסוריים .106 למרות שלא ידוע אם KCNK18 רגיש ישירות לגירוי מכני, הוא עשוי למלא תפקיד בתיווך תגובות למגע קל. כמו גם גירויים מכניים כואבים. KCNK18 ובמידה פחותה KCNK3 מוצעים להיות המטרה המולקולרית של הידרוקסי -? - sanshool, תרכובת שנמצאת בגרגירי פלפל של Schezuan המפעילה קולטני מגע ומעוררת תחושת עקצוץ בבני אדם. 107,108
ערוץ K + KCNQ4 (Kv7.4) תלוי ב- KK4 (KV4) הוא קריטית לקביעת מהירות ההעדפות והתדר של תת-שכבתית של מכניזמים המותאמים במהירות בשני העכברים ובני האדם. מוטציה של KCNQ4 כבר קשורה בתחילה עם סוג של חירשות תורשתית. מעניין מחקר שנערך לאחרונה KCNQ4 בקצות העצבים ההיקפיים של עורית מהירה מסתגלת זקיק השיער ו Meissner גופי. לפיכך, אובדן של הפונקציה KCNQ109 מוביל לשיפור סלקטיבי של רגישות mechanoreceptor לרטט בתדירות נמוכה. יש לציין, אנשים עם אובדן שמיעה מאוחרת עקב מוטציות דומיננטיות של הגן KCNQXNUMX להראות ביצועים משופרים באיתור רטט קטן משרעת, תדר נמוך. XNUMX

תובנה של ד"ר אלכס חימנז
מגע נחשב לאחד החושים המורכבים ביותר בגוף האדם, במיוחד כי אין איבר מסוים אחראי על זה. במקום זאת, תחושת המגע מתרחשת דרך קולטנים סנסוריים, הידועים כ"מכניסטפטורים ", הנמצאים על פני העור ומגיבים ללחץ מכני או לעיוות. ישנם ארבעה סוגים עיקריים של מכאנספטקטורים בעור הירקרק, או חסר השיער, של יונקים: גופי למלר, גופי מישוש, קצות עצבים של מרקל וגופים בולבוסים. מנגנונים פועלים על מנת לאפשר זיהוי של מגע, על מנת לפקח על המיקום של השרירים, העצמות והמפרקים, הידועים בשם proprioception, ואפילו כדי לזהות את הצלילים ואת תנועת הגוף. הבנת מנגנוני המבנה והתפקוד של מכניזמים אלה היא מרכיב בסיסי בשימוש בטיפולים ובטיפולים לניהול כאב.
סיכום
מגע הוא תחושה מורכבת משום שהוא מייצג תכונות מישושיות שונות, כלומר, רטט, צורה, מרקם, הנאה וכאב, עם מופעים שונים מפלים. עד כה, ההתאמה בין איבר מגע לבין התחושה הפסיכו-פיזית הייתה קורלטיבית וסמנים מולקולריים ספציפיים למחלקה רק מתעוררים. פיתוח של בדיקות מכרסם התואמות את מגוון ההתנהגות המגע נדרש כעת כדי להקל על זיהוי גנום בעתיד. השימוש בעכברים ללא תת-קבוצות ספציפיות של סוגי חושים סנסוריים יקל מאוד על זיהוי של מכני-מוחיים וסיבים מודעים חושיים הקשורים במודעת מגע מסוימת. מעניין לציין, כי מאמר שפורסם לאחרונה פותח את השאלה החשובה של הבסיס הגנטי של תכונות מכאוסנסוריות בבני אדם, ומציע כי מוטציה גנטית אחת יכולה להשפיע לרעה על רגישות המגע. 110 זה מדגיש כי הפתופיזיולוגיה של הגירעון מגע אנושי הוא חלק גדול לא ידוע ובוודאי התקדמות על ידי זיהוי מדויק של קבוצת נוירונים סנסוריים המקושרים למגע או לגעת במגע.
בתמורה, נעשתה התקדמות כדי להגדיר את המאפיינים הביופיזיים של הזרמים מכני מגודר. 64 פיתוח של טכניקות חדשות בשנים האחרונות, המאפשר ניטור של שינויים בממברנה במתח, תוך הקלטת מכני מגודר הנוכחי, הוכיח שיטה ניסיונית יקר לתאר זרמים מכאנסוסיטיביים עם הסתגלות מהירה, בינונית ואיטית (שנבדקו בדלמאס ומשתפי פעולה) .XNX העתיד יהיה לקבוע את התפקיד של המאפיינים הנוכחיים במנגנוני הסתגלות של mechanoreceptors מגוונת מבחינה תפקודית ואת התרומה של זרמי K + מכני רגישות לרגישות של LTMRs ו- HTMRs.
האופי המולקולרי של זרמי מכנו-מגודלים ביונקים הוא גם נושא מחקר מבטיח בעתיד. מחקר עתידי יתקדם בשתי נקודות מבט, תחילה כדי לקבוע את התפקיד של מולקולה אביזר כי לקשור ערוצי השלד ואת יידרש כדי להעניק או להסדיר mechanosensitivity של ערוצי יון של כמו TRP ו ASIC / משפחות EnaC. שנית, לחקור את השטח הגדול והמבטיח של התרומה של ערוצי Piezo על ידי מענה לשאלות מפתח, יחסית למנגנוני החדירה והמנגנון, תת-הקבוצות של נוירונים סנסוריים ואופני מגע הקשורים ל- Piezo ותפקיד Piezo בתאי עצב שאינם קשורים מכושנות.
חוש המישוש, בהשוואה לזה של הראייה, הטעם, הצליל והריח, המשתמשים באיברים ספציפיים לעיבוד תחושות אלו, יכול להתרחש בכל הגוף דרך קולטנים זעירים המכונים מכני רצפטורים. ניתן למצוא סוגים שונים של מכני רצפטורים בשכבות שונות של העור, שם הם יכולים לזהות מגוון רחב של גירויים מכניים. המאמר לעיל מתאר דגשים ספציפיים המדגימים את התקדמותם של מנגנונים מבניים ותפקודיים של קולטני מכני הקשורים לחוש המישוש. מידע שהוזכר מהמרכז הלאומי למידע ביוטכנולוגיה (NCBI). היקף המידע שלנו מוגבל לכירופרקטיקה כמו גם לפציעות ולעמוד בעמוד השדרה. כדי לדון בנושא, אל תהסס לשאול את ד"ר חימנז או ליצור איתנו קשר ב 915-850-0900 .
אושר על ידי ד"ר אלכס Jimenez

נושאים נוספים: כאב גב
כאב גב הוא אחד הגורמים הנפוצים ביותר עבור נכות וחסר ימים בעבודה ברחבי העולם. לאמיתו של דבר, כאב גב יוחס כסיבה השנייה הנפוצה ביותר לביקורים של רופא, עולה במספרם רק על ידי זיהומים בדרכי הנשימה העליונות. בערך 80 אחוז מהאוכלוסייה יחוו איזה סוג של כאבי גב לפחות פעם אחת במהלך חייהם. עמוד השדרה הוא מבנה מורכב המורכב מעצמות, מפרקים, מיתרים ושרירים, בין רקמות רכות אחרות. בגלל זה, פציעות ו / או תנאים מחמירים, כגון דיסקים, יכול בסופו של דבר להוביל תסמינים של כאבי גב. פציעות ספורט או פציעות תאונות דרכים הם לעתים קרובות הגורם השכיח ביותר של כאבי גב, עם זאת, לפעמים הפשוטה ביותר של תנועות יכול להיות תוצאות כואבות. למרבה המזל, אפשרויות טיפול אלטרנטיביות, כגון טיפול כירופרקטי, יכולות לעזור להקל על כאבי גב באמצעות שימוש בהתאמות השדרה ומניפולציות ידניות, ובסופו של דבר משפרות את תחושת הכאב.

EXTRA חשוב נושא: כאב גב נמוך ניהול
נושאים נוספים: EXTRA EXTRA: כאבים וטיפולים כרוניים
רֵיק
הפניות
סגור אקורדיון
המידע כאן בנושא "מנגנונים מבניים ופונקציונליים של מכני" אינו מיועד להחליף מערכת יחסים אחד על אחד עם איש מקצוע מוסמך בתחום הבריאות או רופא מורשה ואינו ייעוץ רפואי. אנו ממליצים לך לקבל החלטות בתחום הבריאות על סמך המחקר והשותפות שלך עם איש מקצוע מוסמך.
היקף המידע שלנו מוגבל לכירופרקטיקה, שרירים ושלד, תרופות פיזיות, בריאות, אטיולוגי תורם הפרעות ויסרוסומטיות במצגות קליניות, דינמיקה קלינית של רפלקס סומטויסצרלי, תסביכי תת-לוקסציה, בעיות בריאותיות רגישות ו/או מאמרים, נושאים ודיונים ברפואה תפקודית.
אנו מספקים ומציגים שיתוף פעולה קליני עם מומחים מדיסציפלינות שונות. כל מומחה נשלט על ידי היקף העיסוק המקצועי שלו וסמכות הרישיון שלו. אנו משתמשים בפרוטוקולים פונקציונליים של בריאות ובריאות כדי לטפל ולתמוך בטיפול בפציעות או הפרעות של מערכת השרירים והשלד.
הסרטונים, הפוסטים, הנושאים, הנושאים והתובנות שלנו מכסים עניינים קליניים, נושאים ונושאים הקשורים ותומכים במישרין או בעקיפין בהיקף העיסוק הקליני שלנו.*
משרדנו ניסה באופן סביר לספק ציטוטים תומכים וזיהה את המחקר או המחקרים הרלוונטיים התומכים בפוסטים שלנו. אנו מספקים עותקים של מחקרי מחקר תומכים הזמינים למועצות הרגולטוריות ולציבור על פי בקשה.
אנו מבינים כי אנו מכסים עניינים הדורשים הסבר נוסף כיצד זה עשוי לסייע בתכנית טיפול מסוימת או בפרוטוקול טיפולי; לכן, כדי לדון בהמשך בנושא לעיל, אנא אל תהסס לשאול ד"ר אלכס חימנז, די.סי, או צור איתנו קשר בכתובת 915-850-0900.
אנחנו כאן כדי לעזור לך ולמשפחתך.
ברכות
ד"ר אלכס חימנז זֶרֶם יָשָׁר, MSACP, RN*, CCST, IFMCP*, CIFM*, ATN*
דוא"ל: coach@elpasofunctionalmedicine.com
מורשה כדוקטור לכירופרקטיקה (DC) ב טקסס & ניו מקסיקו*
מס' רישיון טקסס DC TX5807, ניו מקסיקו DC # רישיון NM-DC2182
מורשה כאחות מוסמכת (RN*) in פלורידה
פלורידה רישיון RN רישיון # RN9617241 (מס' שליטה 3558029)
סטטוס קומפקטי: רישיון רב מדינות: מורשה להתאמן ב 40 מדינות*
ד"ר אלכס חימנז DC, MSACP, RN* CIFM*, IFMCP*, ATN*, CCST
כרטיס הביקור הדיגיטלי שלי





