מרפאת גב צוות כירופרקטיקה ורפואה תפקודית מתח חמצוני. עקה חמצונית מוגדרת כהפרעה באיזון בין ייצור חמצן תגובתי (רדיקלים חופשיים) לבין הגנה נוגדת חמצון. במילים אחרות, מדובר בחוסר איזון בין ייצור רדיקלים חופשיים ליכולת של הגוף לסתור או לנקות את ההשפעות המזיקות באמצעות נטרול על ידי נוגדי חמצון. מתח חמצוני מוביל למצבים פתופיזיולוגיים רבים בגוף. אלה כוללים מחלות ניווניות של עצבים, כלומר מחלת פרקינסון, מחלת אלצהיימר, מוטציות גנים, סרטן, תסמונת עייפות כרונית, תסמונת X שביר, הפרעות לב וכלי דם, טרשת עורקים, אי ספיקת לב, התקף לב ומחלות דלקתיות. חמצון מתרחש במספר נסיבות:
התאים משתמשים בגלוקוז כדי לייצר אנרגיה
המערכת החיסונית היא להדוף חיידקים ויצירת דלקת
הגופים לניקוי רעלים, חומרי הדברה ועשן סיגריות
ישנם מיליוני תהליכים המתרחשים בגופנו בכל זמן נתון שיכול לגרום לחמצון. הנה כמה תסמינים:
עייף
אובדן זיכרון או ערפל המוח
כאבי שרירים או מפרקים
קמטים עם שיער אפור
ירידה בראייה
כאבי ראש ורגישות לרעש
רגישות לזיהומים
בחירת מזונות אורגניים והימנעות מרעלים בסביבה שלך עושה הבדל גדול. זה, יחד עם הפחתת הלחץ, יכול להיות מועיל בהפחתת החמצון.
עבור אנשים המעוניינים לשפר את בריאות הלב, האם צריכת שזיפים מיובשים יכולה לעזור לתמוך בבריאות הלב וכלי הדם?
שזיפים מיובשים ובריאות הלב
שזיפים מיובשים, או שזיפים מיובשים, הם פירות עשירים בסיבים שהם צפופים יותר ברכיבים תזונתיים משזיפים טריים ומסייעים לעיכול ולתנועת המעיים. (אלן לוור ואחרים, 2019) מחקר חדש מצביע על כך שהם יכולים להציע יותר מהקלה על עיכול ועצירות, על פי מחקרים חדשים שהוצגו באגודה האמריקאית לתזונה. אכילת שזיפים מיובשים מדי יום יכולה לשפר את רמות הכולסטרול ולהפחית מתח חמצוני ודלקת.
אכילת חמישה עד 10 שזיפים מיובשים ביום עשויה לתמוך בבריאות הלב.
יתרונות לבריאות הלב של צריכה קבועה נראו אצל גברים.
אצל נשים מבוגרות, לאכילה קבועה של שזיפים מיובשים לא הייתה השפעה שלילית על רמות הכולסטרול הכללי, הסוכר בדם והאינסולין.
מחקר אחר מצא שאכילת 50-100 גרם או חמישה עד עשרה שזיפים מיובשים מדי יום קשורה בסיכון מופחת למחלות לב. (מי יאנג הונג וחב', 2021)
המסקנה הייתה ששזיפים מיובשים יכולים לתמוך בבריאות הלב וכלי הדם.
שזיפים מיובשים ושזיפים טריים
למרות שמחקרים העלו כי שזיפים מיובשים יכולים לתמוך בבריאות הלב, זה לא אומר ששזיפים טריים או מיץ שזיפים מיובשים יכולים להציע את אותם היתרונות. עם זאת, אין הרבה מחקרים על היתרונות של שזיפים טריים או מיץ שזיפים, אבל ייתכן שכן. עם זאת, יש צורך במחקר נוסף. שזיפים טריים שיובשו באוויר חם משפרים את הערך התזונתי ואת חיי המדף של הפרי, וזו יכולה להיות הסיבה שהגרסה המיובשת שומרת על יותר חומרים מזינים. (Harjeet Singh Brar et al., 2020)
ייתכן שאנשים יצטרכו לאכול יותר שזיפים כדי לרכוש את אותן היתרונות.
אכילת 5-10 שזיפים מיובשים נראה קל יותר מאשר לנסות להשוות את אותה כמות, או יותר, של שזיפים טריים.
אבל כל אחת מהאפשרויות מומלצת במקום מיץ שזיפים, מכיוון שפירות שלמים מכילים יותר סיבים, גורמים לגוף להרגיש מלא יותר, ובעלי פחות קלוריות.
הטבות לצעירים
רוב המחקר בוצע על נשים וגברים מעל גיל 55 לאחר גיל המעבר, אך אנשים צעירים יותר יכולים להפיק תועלת מאכילת שזיפים מיובשים. תזונה עשירה בפירות וירקות נחשבת לבריאה, ולכן הוספת שזיפים מיובשים לתזונה תוסיף ליתרונות בריאותיים. לאנשים שאינם אוהבים שזיפים מיובשים, פירות כמו תפוחים ופירות יער מומלצים גם לבריאות הלב. עם זאת, פירות מהווים רק חלק אחד בתזונה, וחשוב להתמקד בתזונה מאוזנת עם ירקות, קטניות ושמנים בריאים ללב. שזיפים מיובשים מכילים הרבה סיבים, לכן מומלץ לאנשים להוסיף אותם לאט לשגרת היומיום שלהם, שכן הוספת יותר מדי בבת אחת עלולה להוביל להתכווצויות, נפיחות ו/או עצירות.
כיבוש אי ספיקת לב
הפניות
Lever, E., Scott, S. M., Louis, P., Emery, P. W., & Whelan, K. (2019). השפעת שזיפים מיובשים על תפוקת צואה, זמן מעבר מעיים ומיקרוביוטה של מערכת העיכול: ניסוי אקראי מבוקר. תזונה קלינית (אדינבורג, סקוטלנד), 38(1), 165–173. doi.org/10.1016/j.clnu.2018.01.003
Hong, M. Y., Kern, M., Nakamichi-Lee, M., Abbaspour, N., Ahouraei Far, A., & Hooshmand, S. (2021). צריכת שזיפים מיובשים משפרת את הכולסטרול הכללי ואת קיבולת נוגדי החמצון ומפחיתה דלקת בנשים בריאות לאחר גיל המעבר. כתב עת למזון רפואי, 24(11), 1161–1168. doi.org/10.1089/jmf.2020.0142
Harjeet Singh Brar, Prabhjot Kaur, Jayasankar Subramanian, Gopu R. Nair & Ashutosh Singh (2020) Effect of Chemical Pretreatment on Drying Kinetics and Physio-chemical Characteristics of Yellow European Plums, International Journal of Fruit Science, 20:sup2, S252 , DOI: 279/10.1080
ד"ר אלכס חימנז, DC, מציג כיצד מתח כרוני יכול להשפיע על הגוף וכיצד הוא מתאם עם דלקת בסדרה זו בת שני חלקים. חלק 1 בדק כיצד מתח מתאם עם סימפטומים שונים המשפיעים על רמות הגנים בגוף. חלק 2 בוחן כיצד דלקת ומתח כרוני מתואמים עם הגורמים השונים שיכולים להוביל להתפתחות גופנית. אנו מפנים את המטופלים שלנו לספקים רפואיים מוסמכים המספקים טיפולים זמינים עבור אנשים רבים הסובלים ממתח כרוני הקשור למערכת הלב וכלי הדם, האנדוקרינית והחיסון המשפיעות על הגוף ומפתחות דלקת. אנו מעודדים כל אחד מהמטופלים שלנו על ידי אזכורם בפני ספקים רפואיים קשורים בהתבסס על הניתוח שלהם כראוי. אנו מבינים שחינוך הוא דרך מענגת כששואלים את הספקים שלנו שאלות לפי בקשתו והבנתו של המטופל. ד"ר Jimenez, DC, משתמש במידע זה רק כשירות חינוכי. כתב ויתור
כיצד לחץ יכול להשפיע עלינו?
ד"ר אלכס חימנז, DC, מציג: מתח יכול ליצור רגשות רבים שיכולים להשפיע רבות על רבים מאיתנו. בין אם זה כעס, תסכול או עצב, לחץ יכול לגרום לכל אחד להגיע לנקודת שבירה ולגרום למצבים בסיסיים שעלולים להתפתח לבעיות קרדיווסקולריות. אז לאנשים עם רמת הכעס הגבוהה ביותר, כאשר אתה מסתכל על ספרות הלב וכלי הדם, יש את ההסתברות הנמוכה ביותר לשרוד. כעס הוא שחקן גרוע. כעס גורם להפרעות קצב. המחקר הזה בדק, עכשיו כשיש לנו אנשים עם ICD ודפיברילטורים, אנחנו יכולים לנטר את הדברים האלה. ואנחנו רואים שכעס יכול לעורר הפרעות קצב חדריות בחולים. ועכשיו קל לעקוב אחריו, עם חלק מהטכנולוגיה שלנו.
כעס נקשר לפרקים של פרפור פרוזדורים. כשחושבים על זה, זה אדרנלין שזורם לגוף וגורם להתכווצות כלילית. זה מגביר את קצב הלב. כל הדברים האלה יכולים להוביל להפרעות קצב. וזה לא חייב להיות AFib. זה יכול להיות APCs ו-VPCs. כעת, יצא מחקר מעניין מאוד על טלומראז וטלומרים. טלומרים הם מכסים קטנים על הכרומוזומים, והטלומראז הוא האנזים המקושר ליצירת טלומרים. ועכשיו, אנחנו יכולים להבין דרך שפת המדע, ואנחנו מתחילים להשתמש בטכנולוגיה ולהשתמש במדע באופן שמעולם לא יכולנו לעשות קודם כדי להבין את השפעת הלחץ על טלומרים ואנזימי טלומראז.
הגורמים המובילים ללחץ כרוני
ד"ר אלכס חימנז, DC, מציג: אז אחד מאנשי המפתח לחקור את זה הוא זוכת פרס נובל, ד"ר אליזבת בלקבורן. ומה שהיא אמרה זה שזו מסקנה, ונחזור לכמה ממחקרים אחרים שלה. היא מספרת לנו שהטלומרים של תינוקות מנשים ברחם סבלו מלחץ רב או אפילו היו קצרים יותר בבגרות הצעירה בהשוואה לאמהות שלא סבלו מאותם מצבי לחץ. מתח פסיכולוגי אימהי במהלך ההריון עלול להפעיל אפקט תכנות על מערכת הביולוגיה המתפתחת של הטלומרים, שכבר ניכרת בלידה כפי שהיא משתקפת בהגדרת אורך הטלמטריה של לויקוציטים של יילוד. כך שילדים יכולים להיכנס מוטבעים, וגם אם כן, זה יכול להשתנות.
מה לגבי אפליה גזעית הקופסאות האלה כאן מראות אפליה גזעית גבוהה המובילה לאורך טלומרים נמוך, שרובנו חשבנו עליו אי פעם. אז, אורך טלומרים קצר יותר מוביל לסיכון מוגבר לסרטן ולתמותה כוללת. שיעורי שכיחות הסרטן הם 22.5 לכל 1000 שנות אדם בקבוצת הטלומרים הקצרה ביותר, פסוק 14.2 בקבוצה האמצעית ו-5.1 בקבוצת הטלומרים הארוכה ביותר. טלומרים קצרים יותר יכולים להוביל לאי יציבות של הכרומוזום ולגרום להיווצרות סרטן. אז, עכשיו אנחנו מבינים, דרך שפת המדע, את השפעת הלחץ על האנזים הטלומראז ואורך הטלומרים. לדברי ד"ר אליזבת בלקבורן, 58 נשים לפני גיל המעבר היו מטפלות בילדיהן החולים במחלות כרוניות, נשים שהיו להן ילדים בריאים. הנשים נשאלו כיצד הן תופסות מתח בחייהן והאם הוא משפיע על בריאותן על ידי השפעה על ההזדקנות הסלולרית שלהן.
זו הייתה שאלת המחקר כשהם בחנו את אורך הטלומרים ואת האנזים הטלומראז, וזה מה שהם מצאו. כעת, מילת המפתח כאן נתפסת. אסור לנו לשפוט אחד את הלחץ של זה. לחץ הוא אישי, וחלק מהתגובות שלנו עשויות להיות גנטיות. לדוגמה, מי שיש לו תרופות הומוזיגוטיות עם גן איטי עשוי לסבול מהרבה יותר חרדה מאשר למי שאין לו פולימורפיזם גנטי זה. מישהו שיש לו MAOA ב-MAOB עשוי להיות חרדה יותר מאשר מישהו שאין לו את הפולימורפיזם הגנטי הזה. אז יש מרכיב גנטי בתגובה שלנו, אבל מה שהיא מצאה היה לחץ פסיכולוגי. ומספר שנות הטיפול בילדים חולים כרוניים היה קשור באורך טלומרים קצר יותר ופחות פעילות טלומראז, מה שמספק את האינדיקציה הראשונה לכך שלחץ יכול להשפיע על תחזוקת הטלומרים ועל אריכות החיים.
איך לשנות את תגובת הלחץ שלנו?
ד"ר אלכס חימנז, DC, מציג: זה חזק, וספקי שירותי בריאות רבים נמצאים תחת צורה כלשהי של לחץ. והשאלה היא, מה אנחנו יכולים לעשות כדי לשנות את התגובה שלנו? פרמינגהם גם הסתכל על דיכאון וזיהה דיכאון קליני כסיכון גדול יותר לאירועים קרדיו-וסקולריים ולתוצאות גרועות מאשר עישון, סוכרת, LDL גבוה ו-HDL נמוך, וזה מטורף כי אנחנו מבלים את כל הזמן שלנו בדברים האלה. עם זאת, אנחנו לא משקיעים זמן רב בהתמודדות עם ההיבטים הרגשיים של מחלות כלי דם. זה דיכאון מושפע, מלאי, בדיקת סקר פשוטה לדיכאון, הסתכלות על אנשים עם רמות גבוהות של דיכאון לעומת רמות נמוכות של דיכאון. ואתה יכול לראות שככל שאתה הולך מהרמה הנמוכה לגבוהה ביותר, ככל שאתה מתקדם, הסיכוי לשרוד הולך ופוחת.
ולרבים מאיתנו יש את התיאוריות שלנו באשר למה זה קורה. והאם זה בגלל שאם אנחנו בדיכאון, אנחנו לא אומרים, "אה, אני הולך לאכול כמה נבטים, ואני הולך לקחת ויטמיני B האלה, ואני הולך לצאת להתאמן, ואני הולך לעשות קצת מדיטציה." אז גורם סיכון עצמאי לאחר MI לאירוע הוא דיכאון. הלך הרוח שלנו בנוגע לדיכאון גורם לנו לא להיות מסוגלים לתפקד כרגיל ויכול לגרום לגופנו לפתח בעיות המשפיעות על האיברים החיוניים, השרירים והמפרקים שלנו. אז דיכאון הוא שחקן גדול, שכן 75% ממקרי המוות לאחר MI קשורים לדיכאון, נכון? אז בהסתכלות על חולים, עכשיו, אתה צריך לשאול את השאלה: האם זה הדיכאון שגורם לבעיה, או שזו מחלת הציטוקינים שכבר הובילה למחלת הלב שגורמת לדיכאון? אנחנו צריכים לקחת את כל זה בחשבון.
ומחקר נוסף בדק יותר מ-4,000 אנשים ללא מחלה כלילית בתחילת הדרך. על כל עלייה של חמש נקודות בסולם הדיכאון, זה העלה את הסיכון ב-15%. ואלו עם ציוני הדיכאון הגבוהים ביותר היו בעלי שיעור מחלות עורקים כליליים גבוה ב-40% ושיעור תמותה גבוה ב-60%. אז בעיקר כולם חושבים שזו מחלת ציטוקינים שמובילה ל-MI, מחלות כלי דם ודיכאון. ואז, כמובן, כשיש לך אירוע, ואתה יוצא עם שורה שלמה של נושאים סביבו, אנחנו יודעים שלאנשים שנמצאים בדיכאון יש עלייה של פי שניים בתמותה, עלייה של פי חמישה במוות לאחר התקף לב, ו תוצאות גרועות עם ניתוח. זה ככה, מה בא קודם, התרנגולת או הביצה?
כיצד דיכאון קשור ללחץ כרוני?
ד"ר אלכס חימנז, DC, מציג: כל מנתח יודע זאת. הם לא רוצים לעשות ניתוח באנשים מדוכאים. הם יודעים שהתוצאה לא טובה, וכמובן, יש סיכוי נמוך יותר שהם ימלאו אחר כל ההמלצות הנהדרות שלנו לרפואה תפקודית. אז מה הם חלק מהמנגנונים של תפקוד לקוי אוטונומי הוערכו שונות בקצב הלב ורמות נמוכות של אומגה 3, שיש להם השפעה עמוקה על המוח, ורמות נמוכות של ויטמין D. ישנם ציטוקינים דלקתיים שדיברנו עליהם שאינם מקבלים שינה משקמת, ולרבים מחולי הלב שלנו יש דום נשימה. ותזכור, אל תחשוב שזה רק חולי לב כבדים עם צוואר קצר עבה; זה יכול להיות די מטעה. ובאמת חשוב להסתכל על מבנה הפנים וכמובן הקשר החברתי שהוא הרוטב הסודי. אז האם חוסר תפקוד אוטונומי הוא מנגנון? מחקר אחד בחן את השונות בקצב הלב אצל אנשים עם MI לאחרונה, והם בדקו יותר מ-300 אנשים עם דיכאון ואלה ללא דיכאון. הם מצאו שארבעה מדדי שונות של קצב הלב יורידו באנשים עם דיכאון.
דלקת מעיים ומתח כרוני
ד"ר אלכס חימנז, DC, מציג: אז הנה שתי קבוצות של אנשים הסובלים מהתקף לב ושונות בקצב הלב, העולות לפסגה כאטיולוגיה אפשרית. אחד מהדברים הרבים שיכולים להשפיע גם על מתח כרוני בגוף הוא האופן שבו המיקרוביום של המעי ממלא את תפקידו בלחץ חמצוני. המעי הוא הכל, וחולי לב רבים צוחקים כי הם היו שואלים את הקרדיולוגים שלהם, "למה אכפת לך מהמיקרוביום שלי במעיים? למה שזה ישפיע על הלב שלי?" ובכן, כל הדלקת במעיים גורמת למחלת ציטוקינים. ומה שרבים מאיתנו שכחו מאז הלימודים לרפואה הוא שרבים מהנוירוטרנסמיטורים שלנו מגיעים מהמעיים. אז נראה שדלקת כרונית וחשיפה לציטוקינים דלקתיים מובילות לשינויים בתפקוד הדופמין ובגרעיני הבסיס, המשתקפים בדיכאון, עייפות והאטה פסיכומוטורית. אז לא נוכל להדגיש מספיק את התפקיד של דלקת ודיכאון אם נסתכל על תסמונת כלילית חריפה ודיכאון, שהיו קשורים לסמנים גבוהים יותר לדלקת, CRP גבוה יותר, HS נמוך יותר, שונות דופק נמוכה יותר, ומשהו שמעולם לא נבדקת בבית החולים, כלומר חוסרים תזונתיים.
ובמקרה זה, הם בדקו את רמות האומגה 3 וויטמין D, כך שלכל הפחות, בדיקת אומגה 3 ורמת ויטמין D מוצדקות בכל המטופלים שלנו. ובוודאי, אם אתה יכול לקבל אבחנה מלאה לדלקת הנגרמת על ידי מתח. מצב נוסף שאתה חייב להסתכל עליו כשמדובר בדלקת הנגרמת על ידי מתח הוא אוסטאופורוזיס במפרקים. אנשים רבים עם אוסטיאופורוזיס יסבלו מאובדן שרירים, חוסר תפקוד חיסוני, שומן סביב קו האמצע וסוכר גבוה בדם קשורים להזדקנות, וזה יכול לנבוע מרמות גבוהות של קורטיזול בגוף.
סיכון גבוה למחלות לב לקורטיזול גבוה פי שניים בקרב אנשים הנוטלים מינונים גבוהים של סטרואידים. לכמויות קטנות של סטרואידים אין את אותו סיכון, אז זה לא עניין גדול. כמובן, אנחנו מנסים להוציא מהמטופלים שלנו סטרואידים. אבל הנקודה כאן היא שקורטיזול הוא הורמון סטרס והוא הורמון סטרס שמעלה את לחץ הדם ומעלה משקל על קו האמצע, גורם לנו לסוכרתיים, גורם לתנגודת לאינסולין והרשימה אינסופית. אז, קורטיזול הוא שחקן גדול, וכשזה מגיע לרפואה פונקציונלית, עלינו להסתכל על הבדיקות השונות הנוגעות לרמות גבוהות של קורטיזול כמו רגישות למזון, שסתום צואה ל-3 ימים, שסתום נוטרה ולחץ יותרת הכליה. בדיקת אינדקס כדי לבחון מה קורה עם החולים. כאשר יש מערכת עצבים סימפטית מוגברת וקורטיזול גבוה, דנו בכל דבר, החל מקרישה ועד ירידה בקצב הלב, השמנה מרכזית, סוכרת ויתר לחץ דם.
יחסי הורים ומתח כרוני
ד"ר אלכס חימנז, DC, מציג: והפעלת מערכת הרנין-אנגיוטנסין הכל קשור ללחץ. בואו נסתכל על המחקר הזה שבדק 126 סטודנטים לרפואה בהרווארד, ואחריהם עקבו במשך 35 שנים, מחקר ארוך. והם אמרו, מהי השכיחות של מחלות משמעותיות, מחלות לב, סרטן, יתר לחץ דם? והם שאלו את התלמידים האלה שאלות מאוד פשוטות, מה היו היחסים שלך עם אמא שלך ואבא שלך? זה היה מאוד קרוב? היה חם וידידותי? זה היה סובלני? זה היה מתוח וקר? זה מה שהם מצאו. הם גילו שאם התלמידים זיהו את מערכת היחסים שלהם עם הוריהם כמתוחה 100% שכיחות של סיכון בריאותי משמעותי. שלושים וחמש שנים מאוחר יותר, אם אמרו שזה חם וקרוב, התוצאות הורידו את האחוז הזה בחצי. וזה יעזור אם תחשוב מה זה ומה יכול להסביר את זה, ותראה איך חוויות ילדות שליליות גורמות לנו לחלות תוך כמה דקות ואיך אנחנו לומדים את כישורי ההתמודדות שלנו מההורים שלנו.
סיכום
ד"ר אלכס חימנז, DC, מציג: המסורת הרוחנית שלנו מגיעה מההורים שלנו לעתים קרובות. ההורים שלנו הם אלה שמלמדים אותנו לעתים קרובות איך לכעוס או איך לפתור סכסוכים. אז להורים שלנו הייתה השפעה עמוקה עלינו. וכשחושבים על זה, גם החיבור שלנו לא מאוד מפתיע. זהו מחקר מעקב של 35 שנה.
מתח כרוני יכול להוביל לבעיות מרובות שעלולות להיות קשורות למחלות ותפקוד לקוי בשרירים ובמפרקים. זה יכול להשפיע על מערכת המעיים ולהוביל לדלקת אם לא מטפלים בה באופן מיידי. אז כשזה מגיע להשפעה של מתח המשפיע על חיי היומיום שלנו, זה יכול להיות גורמים רבים, ממצבים כרוניים ועד להיסטוריה משפחתית. אכילת מזונות מזינים עשירים בנוגדי חמצון, פעילות גופנית, תרגול מיינדפולנס והליכה לטיפולים יומיים יכולים להוריד את ההשפעות של מתח כרוני ולהפחית את התסמינים הנלווים שחופפים וגורמים לכאבים בגוף. אנו יכולים להמשיך במסע הבריאות והרווחה שלנו ללא כאבים על ידי שימוש בדרכים שונות להורדת מתח כרוני בגופנו.
ד"ר אלכס חימנז, DC, מציג כיצד מתח יכול להשפיע על אנשים רבים ולהתאם עם מצבים רבים בגוף בסדרה זו בת שני חלקים. אנו מפנים את המטופלים שלנו לספקים רפואיים מוסמכים המספקים מספר טיפולים זמינים עבור אנשים רבים הסובלים מיתר לחץ דם הקשור למערכת הלב וכלי הדם, האנדוקרינית והחיסון המשפיעות על הגוף. אנו מעודדים כל אחד מהמטופלים שלנו על ידי אזכורם בפני ספקים רפואיים קשורים בהתבסס על הניתוח שלהם כראוי. אנו מבינים שחינוך הוא דרך מענגת כששואלים את הספקים שלנו שאלות לפי בקשתו והבנתו של המטופל. ד"ר Jimenez, DC, משתמש במידע זה רק כשירות חינוכי. כתב ויתור
כיצד מתח משפיע על הגוף
ד"ר אלכס חימנז, DC, מציג: כעת כל אחד מגיב לשינויים בסביבה אחרת. כאשר מדובר באנשים רבים שעושים פעילויות יומיומיות החל מעבודה בעבודה שלהם, פתיחה בסופי שבוע, פקקים, ביצוע מבחנים או הכנה לנאום גדול, הגוף עובר מצב קבוע של תגובתיות יתר ועד לשלב של תשישות רגשית ונפשית. מה שמשאיר את הפרט מותש ולחוץ. והמפתח הוא לזהות זאת לפני שזה קורה, מכיוון שאנו רואים את ההשפעה הזו של לחץ על המטופלים שלנו ועל עצמנו. והדבר הראשון שצריך להבין הוא מה האירוע היוזם גורם להשפעה הזו.
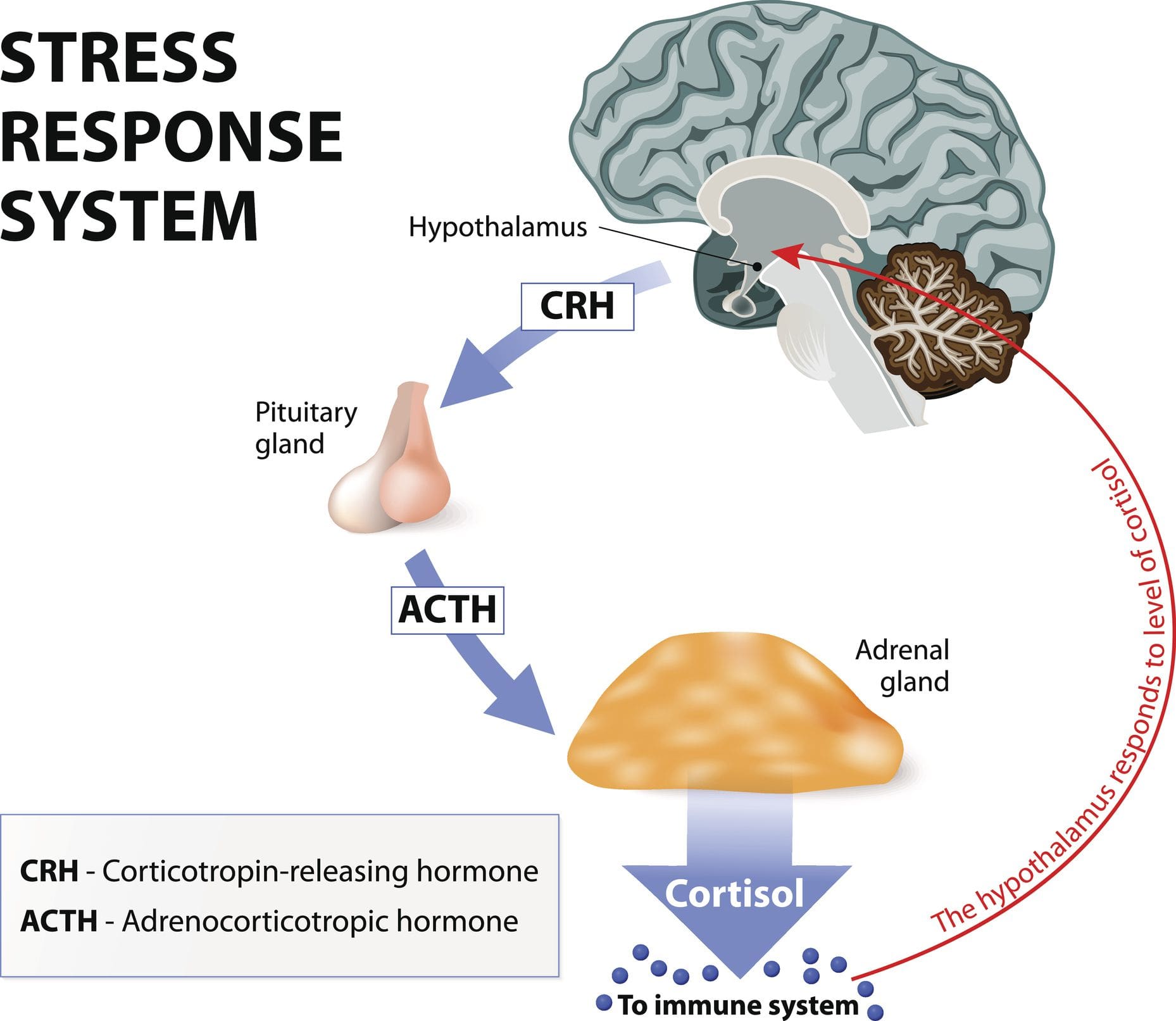
לא משנה מה האירוע היוזם, החלק החשוב ביותר הוא התפיסה שלנו לגבי האירוע. מה זה אומר לנו? האם זו התפיסה שלנו? כאשר הגוף עובר אירוע יוזם זה, זה יכול לגרום לתפיסה להוביל לתגובה ולהשפעה על הגוף שלנו. אז תפיסה היא הכל כשאנחנו מדברים על מתח ותגובת הלחץ. כעת, יש לנו למעלה מ-1400 תגובות כימיות המתרחשות בגוף. אז לצורך ההרצאה הזו, נדון בשלושת המפתחות: אדרנלין ונוירו-אדרנלין, אלדוסטרון וכמובן קורטיזול.
ולמה אלו חשובים? כי לכל אחד מאלה יש השפעה עצומה על מחלות לב וכלי דם. כעת, בשנות ה-1990, רופאים רבים החלו להבין את השפעת הלחץ על הגוף הפיזי. ומה קורה לאנשים כאשר ציר ה-HPA שלהם מאותת שהם נמצאים תחת איום ומתחילים להציף את גופם בהורמוני לחץ? ובכן, אנו רואים קרישה משופרת. אנו רואים שינוי במערכת הרנין והאנגיוטנסין. זה עולה בסיבובים. אנו רואים עלייה במשקל אצל אנשים ותנגודת לאינסולין. מה שהרבה אנשים לא מבינים הוא שהשומנים הופכים לא נורמליים עם מתח. כמעט כל אחד מהמטופלים שלנו יודע שטכיקרדיה והפרעות קצב מתרחשות כאשר האדרנלין שלנו זורם, ולחץ הדם שלנו עולה. עכשיו, חשבו על זה דרך שפת הרפואה.
בסביבות שנות ה-1990, רופאים נתנו אז אספירין ופלביקס לצורך קרישה. אנו ממשיכים לספק ACEs ו-ARBs למטופלים שלנו. השפעת הקורטיזול גורמת לעלייה במשקל ולתנגודת לאינסולין. אנחנו נותנים סטטינים; אנחנו נותנים מטפורמין. אנו מספקים חוסמי בטא עבור זה, טכיקרדיה וחוסמי סידן עבור לחץ דם גבוה זה. אז כל הורמון שנדלק מלחץ, יש לנו תרופה שאנחנו משתמשים בה כדי לאזן את זה. ולמען האמת, במשך שנים, דיברנו על כמה חוסמי בטא טובים ללב. ובכן, כשחושבים על זה, חוסמי בטא אכן חוסמים אדרנלין. אז כשרופאים מסתכלים על זה, הם מתחילים לחשוב, "טוב, אולי אנחנו צריכים לעשות תרופות ולעשות מדיטציה, נכון? אנחנו משתמשים בכל התרופות האלה, אבל אולי נצטרך לבחון דרכים אחרות לשנות את תגובת הלחץ".
מהי כיווץ כלי דם?
ד"ר אלכס חימנז, DC, מציג: לא נקרא כל אחד מהתסמינים האלה כי יש כל כך הרבה, אבל הכל מסתכם באותו דבר. לחץ. אנחנו צריכים לחשוב על מישהו שעבר תאונת דרכים, למשל, והאדם הזה מדמם. אז הגוף יפה בכך שהוא מרכיב דרך למנוע מהאדם דימום או התכווצות כלי דם. כיווץ כלי דם בונה את כלי הדם הללו והופכת את טסיות הדם לדביקות כך שהן יוצרות קריש, והדם יכול להפסיק. זה מגביר את תפוקת הלב על ידי העלאת קצב הלב ומגביר את האלדוסטרון, מה שגורם לאגירת מלח ומים להעלות את לחץ הדם. אז עבור מישהו במצב חירום רפואי, כמו תאונה, דימום או איבוד נפח, זה היופי של גוף האדם. אבל למרבה הצער, אנו רואים אנשים חיים כך, ממש 24/7. אז אנחנו מכירים את התכווצות כלי הדם ואת דביקות הטסיות, ורואים עלייה בסמנים לדלקת, הומוציסטאין, CRP ופיברינוגן, כולם מגבירים את הסיכון הקרדיווסקולרי.
אנו רואים את ההשפעה של קורטיזול, לא רק מעלה את לחץ הדם, לא רק גורם לסוכרת ולתנגודת לאינסולין, אלא גם הפקדת שומן בטני סביב קו האמצע. ואז, כפי שתראו בעוד מספר דקות, ישנם קשרים בין אירועים מלחיצים לבין הפרעות קצב כמו פרפור פרוזדורים ואפילו פרפור חדרים. לראשונה ברפואה, בקרדיולוגיה, יש לנו תסמונת הנקראת takosubo cardiomyopathy, הנקראת בחיבה תסמונת הלב השבור. וזוהי תסמונת שבה שריר הלב הופך המום בצורה חריפה עד כדי גרימת תפקוד חמור של החדר השמאלי או תפקוד לקוי. ובדרך כלל, זה מופעל על ידי חדשות רעות ואירוע מלחיץ רגשית. זה נראה כאילו מישהו צריך השתלת לב. אז כשאנו חושבים על גורמי הסיכון הישנים של פרמינגהם, אנו אומרים, אילו מהם מושפעים מלחץ?
סימפטומים של מתח
ד"ר אלכס חימנז, DC, מציג: לאנשים יש כל מיני התנהגויות לא מסתגלות להלחיץ, בין אם 20 חברים בחפיסת הסיגריות הזו, אוכלים את הקינבון הזה כי זה עושה לי טוב כרגע, או שכל הקורטיזול יגרום לי להשמין ולסוכרת. שומנים עולים תחת לחץ; לחץ הדם עולה תחת לחץ. אז כל אחד מגורמי הסיכון הללו מושפע מהורמוני לחץ. וכמובן, אנו יודעים שעם הפעלת מערכת ה-RAS או מערכת הרנין-אנגיוטנסין, אנו תמיד רואים החמרה באי ספיקת לב. וזה מתואר מאוד בספרות. וגם, לאלו מכם שעשויים לעבוד בחדר המיון, שאלו את המטופלים שלכם מה הם עשו לפני שהגיעו עם האפיזודה שלהם של אי ספיקת לב או כאבים בחזה. ואתה תשמע סיפורים כמו, צפיתי בסרט גרוע, או צפיתי בסרט מלחמה, או התעצבנתי בגלל משחק הכדורגל, או משהו כזה.
נדבר על שונות בקצב הלב, המושפעת מלחץ. וכמובן, מתח משפיע על היכולת שלנו להתנגד לזיהומים. ואנחנו יודעים שאנשים נמצאים בלחץ כשהם מחוסנים. לדוגמה, לייזרים של Cleco פועלים אך אינם מייצרים נוגדנים לחיסון כשהם במתח. וכמובן, כפי שתראו בעוד דקה, לחץ חמור עלול לגרום למוות לבבי פתאומי, MI וכו'. אז זה שחקן רע שמתעלמים ממנו. ועבור רבים מהמטופלים שלנו, הלחץ מניע את הרכבת. אז כשאנחנו מדברים על אכילת נבטים וכרובית, ואתה יודע, הרבה ירקות עלים ירוקים, ומישהו נמצא בלחץ כל כך שהוא מנסה להבין, "איך אני אעבור את היום? ” הם לא שומעים אף אחד מהדברים האחרים שאנחנו ממליצים עליהם.
לכן, מתח כרוני והפרעות רגשיות, בין אם דיכאון, חרדה או פאניקה, שמים את הרגל שלנו על דוושת הגז ומגבירים את מערכת העצבים הסימפתטית. אנו יודעים שאותם דברים שאנו רואים בהזדקנות, כפי שתראו בעוד דקה, קשורים לרמות מוגברות של הורמוני לחץ, במיוחד קורטיזול. אז בין אם זה אוסטיאופורוזיס, ירידה בצפיפות העצם, הפרעה בתפקוד האנדותל, הפעלת טסיות דם, יתר לחץ דם, השמנה מרכזית או תנגודת לאינסולין, זה נובע מתגובת לחץ. וצריך להיות לנו תוכנית למטופלים שלנו כיצד לטפל בזה. המכון האמריקאי ללחץ אומר ש-75 עד 90% מכלל הביקורים אצל ספקי שירותי בריאות נובעים מהפרעות הקשורות ללחץ. וזה הרבה יותר מדי גבוה, אבל על ידי התבוננות בחולים ולאן הם נכנסו, הם מספרים את סיפוריהם לרופאים שלהם. התוצאות זהות; זה לא משנה אם זה היה כאבי ראש, מתח שרירים, אנגינה, הפרעות קצב או מעי רגיז; כמעט תמיד היה בו גורם למתח כלשהו.
מתח אקוטי וכרוני
ד"ר אלכס חימנז, DC, מציג: יש הבדל בין מתח אקוטי לכרוני עם התפיסה והקשר החברתי שלנו. למרות שאנו צוברים קצת כוח מכוח עליון, לחץ יכול להשפיע על כל אחד, וייתכן שרובנו לא נוכל להתמודד איתו היטב. אז מחקר גדול נעשה לפני שנים רבות על ידי ד"ר ריי והולמס שקבע, לפני 50 שנה, הרכיב שיטה לכימות אירועים משנים חיים. אז בואו נסתכל על כמה תחומים, כמו אירועים משנים חיים. כיצד מתרחשים אירועים משנים חיים וכיצד הם מדורגים? מי הם הגדולים, ומי הם הקטנים?
ואיך הדירוג הזה מוביל לבעיות רפואיות גדולות כמו סרטן, התקף לב ומוות פתאומי בעתיד? אז הם בחנו 43 אירועים משנים חיים, דירגו אותם במקור ודירגו אותם מחדש בשנות התשעים. וחלקם נשארו אותו הדבר. הם נתנו ציון התאמה לאירוע, ואז הם בדקו מספרים שיהיו קשורים למחלה קשה. כך, למשל, אירוע משנה חיים. מספר אחד, 1990 יחידות משנות חיים, הוא מוות של בן זוג. כל אחד יכול היה להתייחס לזה. הגירושים היו מספר שתיים, פרידה מספר שלוש, וסופו של בן משפחה קרוב. אבל גם שמתי לב שחלק מהדברים קיבלו דירוג שהם, אולי לא משתווים, להיות אירוע משנה חיים שיכול להשפיע על תגובת לחץ כמו נישואים או פרישה.
סיכום
ד"ר אלכס חימנז, DC, מציג: אז זה לא היה האירוע היחיד שעשה את ההבדל. זה היה חיבור של אירועים. ומה שהם מצאו לאחר שבדקו 67 רופאים זה אם היה לך ציון יחידה משנה חיים של איפשהו בין אפס ל-50 אחד, לא עניין גדול, אין מחלה רצינית אמיתית, אבל ברגע שהגעת לרף ה-300 הזה, היה 50% סיכוי למחלה קשה. אז ציר הזמן הזה של אירועים בחיי המטופל. אנחנו רוצים לדעת מה קרה בחייהם כשהתסמינים שלהם התחילו ואז להחזיר את זה מוקדם יותר כדי להבין את הסביבה שבה חי האדם הזה. ההשפעה של מתח יכולה לגרום לאנשים רבים לפתח מצבים כרוניים ולהסוות תסמינים אחרים שעלולים להוביל לכאבי שרירים ומפרקים. בחלק 2, נצלול יותר על האופן שבו השפעת הלחץ משפיעה על הגוף והבריאות של האדם.
ד"ר אלכס חימנז, DC, מציג כיצד יתר לחץ דם משפיע על גוף האדם וכמה גורמים שיכולים להגביר יתר לחץ דם אצל אנשים רבים בסדרה זו בת שני חלקים. אנו מפנים את המטופלים שלנו לספקים רפואיים מוסמכים המספקים מספר טיפולים זמינים עבור אנשים רבים הסובלים מיתר לחץ דם הקשור למערכת הלב וכלי הדם והחיסון המשפיעים על הגוף. אנו מעודדים כל אחד מהמטופלים שלנו על ידי אזכורם בפני ספקים רפואיים קשורים בהתבסס על הניתוח שלהם כראוי. אנו מבינים שחינוך הוא דרך מענגת כששואלים את הספקים שלנו שאלות לפי בקשתו והבנתו של המטופל. ד"ר חימנז, DC, משתמש במידע זה רק כשירות חינוכי. כתב ויתור
כיצד לחפש יתר לחץ דם
ד"ר אלכס חימנז, DC, מציג: בואו נחזור לעץ ההחלטות כדי שתוכלו להתחיל לחשוב כיצד תחילו את מודל ה- go-to-it ברפואה פונקציונלית ליתר לחץ דם וכיצד תתחילו להעריך טוב יותר מישהו עם יתר לחץ דם במקום לומר לו שלחץ הדם שלו מוגבר. . האם הגוף מושפע מדלקת, מתח חמצוני או תגובה חיסונית? האם זה משפיע על תפקוד האנדותל או השריר החלק של כלי הדם משלוש הקטגוריות הללו של תגובות, דלקת, מתח חמצוני או תגובה חיסונית? האם אנו בוחרים חוסם תעלות סידן משתן או מעכב ACE? וכדי לעשות את זה, זה ממש חשוב בקטע האיסוף שלנו. אם לוקחים את ההיסטוריה הרפואית וציר הזמן של יתר לחץ הדם שלהם, אתה מקבל רמז לגבי הנזק לאיברים בשאלונים. אתה מסתכל על האנתרופומטריה שלהם.
זה כולל את השאלות הבאות:
מהם הסמנים הדלקתיים?
מהם הסמנים הביולוגיים והאינדיקטורים הקליניים?
אלה מתוארים דרך עץ ההחלטות הקליניות. וכבר רק עשית את זה, אתה הולך להרחיב ולכוון את העדשה שלך לפי מה שאתה עשוי לראות בחולה עם יתר לחץ דם. בואו נוסיף לציר הזמן מתי מתחיל יתר לחץ דם? מסגרת הזמן של יתר לחץ דם מתחילה למעשה לפני הלידה. חשוב לשאול את המטופל שלך אם הם היו בגיל חינוך מוקדם או גבוה. האם אמם הייתה לחוצה? האם הם נולדו מוקדם או פג? האם היה לחץ תזונתי בהריון שלהם? אם הם יודעים את זה, אתה יכול להיות שני אנשים עם אותו גודל כליות, אבל מי שלא היה לו מספיק חלבון במהלך ההריון יכול לסבול עד 40% פחות גלומרולי. הידיעה שזה ישנה את האופן שבו תתאים את התרופה עשרות שנים מאוחר יותר אם אתה יודע שיש להם אולי 40% פחות גלומרולי.
ציר הזמן ללחץ דם
ד"ר אלכס חימנז, DC, מציג: אז חשוב לקחת את ציר הזמן של לחץ הדם שלהם. אז חשוב גם לזהות מה קורה כשאנחנו מתחילים לארגן ולאסוף נתונים דרך הסמנים הביולוגיים; הסמנים הביולוגיים הבסיסיים יתנו לך רמזים לגבי האם יש להם בעיות עם שומנים באינסולין, האם יש להם בעיות בתגובתיות של כלי הדם, איזון מערכת העצבים האוטונומית, חוסר איזון, קרישה או השפעות רעל חיסוני. אז זה דבר הגיוני להדפסה מכיוון שבמטופל שלך עם יתר לחץ דם, זה רק באמצעות הסמנים הביולוגיים אתה יכול להתחיל להבין אילו אזורים של תפקוד לקוי משפיעים על דלקת, מתח חמצוני ותגובה חיסונית וכיצד הסמנים הביולוגיים האלה משקפים את זה מידע עבורך. זה מאוד סביר שיש לפניך כדי לעזור לשנות את המחשבות שלך על יתר לחץ דם וגם מאפשר לך לחדד כמה מהמאפיינים של האדם בצד השני של הסטטוסקופ שלך בצורה אישית ומדויקת יותר.
אבל בואו נתחיל ממש בהתחלה. האם למטופל שלך יש לחץ דם גבוה? אנו יודעים שבהתאם להשפעות איברי הקצה של התחלואה הנלוות שלהם, אתה עלול להפעיל לחץ דם גבוה במקצת אם יש לך בעיית ריבוי במוח ובכליות או בלב, אבל יש כמה קווים מנחים. הנחיות איגוד הלב האמריקאי לשנת 2017 לקטגוריות לחץ דם מפורטות כאן. הם הלכו ודעכו קדימה ואחורה במהלך העשורים האחרונים, אבל זה מאוד ברור. לאחר עליית לחץ דם, כל דבר מעל 120, באמת שינתה את מספר האנשים שאנו מתחילים לראות או שוקלים לטפל בגורמים השורשיים ללחץ הדם שלהם. אז נחזור לזה, במיוחד במקרה כדי לעזור לנו להסתכל על איך אנחנו מסווגים אנשים עם בעיות לחץ דם.
הקריטריונים למדידת לחץ דם
ד"ר אלכס חימנז, DC, מציג: מה הצעד הראשון? זה איך אתה לוקח את לחץ הדם במטופל שלך? האם הם מפקחים על זה בבית? האם הם מביאים לך את המספרים האלה? איך אתה עוקב אחר לחץ הדם במרפאה שלך? איך אתה מקבל קריאות מדויקות במרפאה שלך? להלן הקריטריונים למדידת לחץ דם מדויקת והשאלות שיש לשקול אם אתה עושה את כל אלה.
האם אתה שואל את המטופל שלך אם הוא שתה קפאין בשעה האחרונה?
האם הם עישנו בשעה הקודמת?
האם הם נחשפו לעשן בשעה האחרונה?
האם המקום שבו אתה מוריד לחץ דם חם ושקט?
האם הם יושבים עם גב נתמך בכיסא עם הרגליים על הקרקע?
האם אתה משתמש בשולחן הצד המתגלגל כדי להניח את זרועך בגובה הלב?
האם הם יושבים ליד שולחן הבחינה כשרגליים משתלשלות, וסייעת אחות מרימה את זרועם ומכניסה את קפל בית השחי כדי להחזיק את זרועם שם?
האם רגליהם על הקרקע?
האם הם ישבו שם חמש דקות?
האם הם התאמנו ב-30 הדקות הקודמות?
ייתכן שיש לך לחץ דם סיסטולי אם הכל בקריטריונים. הנה האתגר. ישנם 10 עד 15 מילימטרים של כספית גבוהים יותר כשמדובר בישיבה ונטילת לחץ דם. מה לגבי גודל השרוול? אנחנו מכירים את המאה שעברה; לרוב המבוגרים היה היקף זרוע עליונה של פחות מ-33 סנטימטרים. ליותר מ-61% מהאנשים יש כעת היקף זרוע עליון גדול מ-33 סנטימטרים. אז גודל השרוול שונה עבור כ-60% מהמטופלים המבוגרים שלך, תלוי באוכלוסייתך. אז אתה צריך להשתמש בחפת גדול. אז תסתכל כיצד נאסף לחץ הדם במשרד שלך. נניח שלחץ הדם גבוה אצל המטופלים שלך; אז אנחנו צריכים לשאול, האם זה נורמלי? גדול.
הסוגים השונים של יתר לחץ דם
ד"ר אלכס חימנז, DC, מציג: האם זה מוגבר בגלל יתר לחץ דם במעיל לבן? האם יש להם לחץ דם תקין, מוגבר מחוץ למרפאה, או יתר לחץ דם מוסווה? או שסתם יש להם יתר לחץ דם מתמשך וזה אתגר? על זה נדבר. אז כשאתה מפרש, חשוב לשקול גם ניטור לחץ דם אמבולטורי. אז אם יש לך מישהו עם יתר לחץ דם ואינך יודע אם לחץ הדם יורד ואתה מנסה להבין אם יש לו יתר לחץ דם מתמשך, אתה יכול להשתמש בניטור לחץ דם 24 שעות ביממה. לחץ הדם הממוצע בשעות היום מעל 130 מעל 80 הוא יתר לחץ דם, לחץ הדם הממוצע בלילה מעל 110 מעל 65 הוא יתר לחץ דם. אז למה זה חשוב? לחץ הדם הממוצע יורד לסביבות 15% בלילה בגלל הבעיה עם טבילת לחץ הדם. אי ירידה בלחץ הדם בזמן שאתה ישן בלילה עלול לפתח בעיות שעלולות להשפיע על אדם במהלך היום.
אם המטופל שלך ישן בלילה, הוא אמור לרדת בכ-15% כשהוא ישן. אם יש להם לחץ דם ללא טבילה, זה קשור למחלות נלוות. מהן כמה מאותן מחלות נלוות בלחץ דם ללא טבילה? חלק מהמצבים המתואמים עם לחץ דם ללא טבילה כוללים:
מחלת לב גדושה
מחלות לב וכלי דם
מחלת כלי דם במוח
אי ספיקת לב
כשל כלייתי כרוני
הפרות מוחיות שקטות
מחלות נלוות הקשורות ללא לחץ דם
ד"ר אלכס חימנז, DC, מציג: אלו הן המחלות הנלוות הקשורות ללא לחץ דם. כולנו מסכימים שלחץ דם מוגבר אינו בהכרח טוב בכל אותם תנאים. אז כאשר אתה מסתכל על קבוצות אנשים שונות או מחלות נלוות אחרות, לחץ דם ללא טבילה קשור לרוב לאנשים הרגישים לנתרן, אנשים שיש להם אי ספיקת כליות, אנשים שיש להם סוכרת, אנשים שעברו היפרטרופיה של חדר שמאל, אנשים שיש להם יתר לחץ דם עקשן. או תפקוד לקוי של מערכת העצבים האוטונומית ולבסוף, דום נשימה בשינה. אז, לחץ דם ללא טבילה מגביר את הקשר שלך לנזק לבבי תת-קליני. אוקיי, טבילה הפוכה פירושה שאתה יותר לחץ דם בלילה וקשורה יותר לעלייה מאשר במהלך היום קשורה יותר לשבץ דימומי. ואם יש לך מישהו עם יתר לחץ דם לילי, אתה צריך להתחיל לחשוב על דברים כמו עורקי הצוואר ועלייה בעובי המדיאלי הפנימי של הצוואר. אתה מתחיל לחשוב על היפרטרופיה של חדר שמאל ואולי תראה את זה ב-EKG. הנה מה שאנחנו יודעים על יתר לחץ דם לילי. יתר לחץ דם לילי הוא לחץ דם לילי גבוה מ-120 מעל 70. הוא קשור ליכולת חיזוי רבה יותר של תחלואה ותמותה קרדיווסקולרית.
אם יש לך יתר לחץ דם לילי, זה מעלה את הסיכון לתמותה ממחלות לב וכלי דם ב-29 עד 38%. אנחנו חייבים לדעת מה קורה בלילה כשאנחנו ישנים, נכון? ובכן, מה עוד חידוד? חידוד נוסף הוא ההכרה שלחץ הדם במנוחה נשלט על ידי מערכת הרנין-אנגיוטנסין שלך. לחץ הדם הערות נשלט על ידי מערכת העצבים הסימפתטית שלך. אז בואו נדבר על איך מערכת האנגיוטנסין הכלייתית שלהם מניעה את יתר לחץ הדם הלילי שלהם, ואתם חושבים על איזו תרופה הם לוקחים. ייתכן שתשנה את מינון התרופה ללילה. ובכן, מחקרים הראו שאם יש לך יתר לחץ דם לילי ואתה לא טובל, עדיף ליטול את מעכבי ACE, ARBs, חוסמי תעלות סידן וחוסמי בטא מסוימים בלילה לפני השינה. אבל זה הגיוני שלא תעביר את המשתנים שלך לשעות הלילה, אחרת תהיה לך שינה משבשת.
טיפול בלחץ הדם בשעות היום והלילה
ד"ר אלכס חימנז, DC, מציג: אז אם אנחנו לא מתייחסים ללחץ הדם בשעות היום והלילה, עלינו לשקול את ההשפעה של עומס לחץ הדם. מהו לחץ הדם הממוצע שלך בשעות היום ולחץ הדם המתון שלך בשינה. אנו יודעים שעומס לחץ הדם אצל מבוגרים צעירים הוא יתר לחץ דם רק כ-9% מהמקרים. אז כלומר העומס הסיסטולי הוא כ-9% לעומת קשישים, כ-80% מעומס לחץ הדם הוא סיסטולי. וכך כאשר יש לך עומס סיסטולי גבוה יותר, יש לך יותר סיבוכים ונזק לאיבר הקצה. אז מה שאנחנו מדברים עליו הוא לעזור לזהות את המטופל שלך עם יתר לחץ דם; מה ציר הזמן שלהם מה הפנוטיפ שלהם? האם הם בעלי יתר לחץ דם רק במהלך היום, או שהם בעלי יתר לחץ דם גם בלילה? אנחנו צריכים לבדוק מה עוזר לאזן את זה.
הנה הנקודה השנייה, רק לכ-3.5% מהאנשים עם יתר לחץ דם יש לזה סיבה גנטית. רק 3.5% מהאנשים הגנים שלהם גורמים ליתר לחץ דם. הכוח נמצא בתחתית המטריצה ומזהה את הדפוסים האלה, נכון? אז אתה מסתכל על פעילות גופנית, שינה, דיאטה, מתח ומערכות יחסים. אז אנחנו יודעים שארבעת האיזונים האוטונומיים האלה עוזרים לקבוע את לחץ הדם. נבחן את מערכת האנגיוטנסין הכלייתית, נפח הפלזמה שבו הם אוחזים בכמות גדולה מדי של נוזלים, עומס מלחים משני והפרעה בתפקוד האנדותל. חריגות בכל אחד מאלה עלולות להוביל ליתר לחץ דם. דיברנו על עוד אחד שיכול להוביל ליתר לחץ דם: הקשר בין תנגודת לאינסולין ליתר לחץ דם.
זה נותן לך באופן דיאגרמטי מושג על האינטראקציות הפיזיולוגיות בין תנגודת לאינסולין ליתר לחץ דם. זה משפיע על הגברת הטונוס הסימפתטי והגברת איזון מערכת הכליות-אנגיוטנסין. אז בואו נבלה כמה דקות על מסלול מערכת רנין-אנגיוטנסין אנגיוטנסין עד אנגיוטנסין שני. אנו מנצלים את היתרונות של אנזימים אלה על ידי מתן מעכבים לאנזימים הממירים אנגיוטנסין בחולים עם יתר לחץ דם. עלייה באנגיוטנסין XNUMX מובילה להיפרטרופיה קרדיווסקולרית, מובילה להתכווצות פאזה סימפטטית, נפח דם מוגבר, נוזל נתרן, אצירה ושחרור אלדוסטרון. האם אתה יכול לברר לגבי הסמנים הביולוגיים של המטופלים שלך? האם אתה יכול לשאול האם יש להם רמות רנין גבוהות?
חפש את הסימנים
ד"ר אלכס חימנז, DC, מציג: ובכן, אתה יכול. ניתן לבדוק פעילות רנין בפלזמה ורמות אלדוסטרון. חשוב לעשות זאת אם המטופל שלך סובל מלחץ דם גבוה ומעולם לא קיבל תרופות כי זה המקום שבו תחמוצת החנקן היא כל כך חשובה. זה המקום שבו סינתאז תחמוצת החנקן האנדותל שלך נמצא. זה המקום שבו יש לך מתח עצום והמודינמי. זה המקום שבו צריכה תזונתית של ארגינין או הסביבה המשפיעה על תחמוצת החנקן ממלאת תפקיד כזה בבריאותה של שכבת האנדותליה הזו. אם תחבר את הכל איכשהו, בנס, או לפחות בעיני רוחך, זה יכסה שישה מגרשי טניס אצל מבוגר ממוצע. זה שטח פנים עצום. והדברים שגורמים להפרעה בתפקוד האנדותל אינם חדשות חדשות לאנשים ברפואה תפקודית. לחץ חמצוני מוגבר ודלקת הם שני דברים שהזכרנו שמשפיעים.
ואז, תסתכל על כמה מהמרכיבים האחרים האלה, ה-ADMA שלך מוגבר ומתאם עם תנגודת לאינסולין. הכל מתחיל להיווצר יחד במטריצה שמקיימת אינטראקציה. אז אתה מסתכל על תחלואה נלווית אחת בתסמונת קרדיומטבולית, והיא משפיעה על תחלואה נלווית אחרת. פתאום אתה רואה את הקשר ביניהם או היפרהומוציסטינמיה, שהוא סמן מטבוליזם של פחמן אחד, כלומר אתה בוחן את ההתאמה של חומצה פולית, b12, b6, ריבופלבין, ואת הפעילות הזו של חילוף החומרים הפחמן אחד שלך. אז בואו נסתכל על כמה מסמני הסיכון המתעוררים הללו לשיפור ומעקב בחולים עם יתר לחץ דם. בואו ננתח מחדש את ADMA שוב. ADMA מייצג דימתיל ארגינין אסימטרי. אסימטרי, דימתיל ארגינין הוא סמן ביולוגי של תפקוד לקוי של האנדותל. מולקולה זו מעכבת סינתאז תחמוצת החנקן תוך פגיעה בתפקוד האנדותל, ובכל המחלות הנלוות הקשורות לתסמונת קרדיומטבולית, ADMA יכולה להיות מוגברת.
סיכום
אז, כסקירה מהירה, L-arginine הופך לחנקן תחמוצת באמצעות סינתאז תחמוצת החנקן, והתאמה של תחמוצת החנקן מובילה להרחבת כלי הדם. ADMA חוסמת המרה זו. ואם רמות ה-ADMA שלך מוגברות ורמות תחמוצת החנקן שלך נמוכות, אז יש לך ירידה בהצטברות טסיות תחמוצת החנקן בחמצון LDL. כל כך הרבה דברים מפחיתים תחמוצת החנקן או קשורים לרמות נמוכות יותר של תחמוצת החנקן, דום נשימה בשינה, ארגינין נמוך בתזונה, חלבון, אי ספיקה של אבץ ועישון.
כולם מתמודדים עם לחץ בשלב מסוים בחייהם. בין אם זה יהיה ראיון עבודה, דד-ליין ענק, פרויקט או אפילו מבחן, לחץ קיים כדי לשמור על תפקוד הגוף בכל תרחיש שהגוף עובר. מתח יכול לעזור לווסת את הגוף המערכת החיסונית ועזרה מטבוליזם הומאוסטזיס כאשר הגוף מגביר את האנרגיה שלו במהלך היום. כאשר מתמודדים עם מתח כרוני יכול לגרום להפרעות בתפקוד מטבולי בגוף כמו הפרעות במעיים, דלקת, ועלייה ברמות הגלוקוז בדם. מתח כרוני יכול להשפיע גם על מצב הרוח והבריאות של האדם, הרגלי האכילה ואיכות השינה. המאמר של היום יבדוק אם מתח הוא דבר טוב או דבר רע, כיצד הוא משפיע על הגוף, וההשפעות של מה שמתח כרוני עושה לגוף. הפנה מטופלים לספקים מוסמכים ומיומנים המתמחים בטיפולי מעיים לאנשים הסובלים מנוירופתיה אוטונומית. אנו מנחים את המטופלים שלנו על ידי הפניה לספקים הרפואיים הקשורים לנו בהתבסס על בדיקתם כאשר הדבר מתאים. אנו מוצאים שחינוך הוא קריטי לשאילת שאלות בעלות תובנות לספקים שלנו. ד"ר אלכס Jimenez DC מספק מידע זה כשירות חינוכי בלבד. כתב ויתור
האם אתה מרגיש חרדה כל הזמן? מה דעתך להרגיש כאבי ראש שהם כל הזמן מטרד? מרגישים מוצפים ומאבדים מיקוד או מוטיבציה? כל הסימנים הללו הם מצבי לחץ שאדם עובר. מחקרים הגדירו מתח או קורטיזול כהורמון הגוף המספק מגוון השפעות על תפקודים שונים בכל מערכת. קורטיזול הוא הגלוקוקורטיקואיד העיקרי שמגיע מקליפת יותרת הכליה. במקביל, ציר HPA (היפותלמוס-יותרת המוח-אדרנל) עוזר לווסת את הייצור וההפרשה של הורמון זה לשאר הגוף. כעת קורטיזול יכול להיות מועיל ומזיק לגוף, בהתאם למצב בו האדם נמצא. מחקרים נוספים הזכירו שקורטיזול מתחיל ומשפיע על המוח ועל שאר הגוף שכן מתח בצורתו החריפה יכול לגרום לגוף להסתגל ולשרוד. התגובות החריפות של קורטיזול מאפשרות תפקוד עצבי, קרדיווסקולרי, חיסוני ומטבולי בגוף.
איך זה משפיע על חילוף החומרים של הגוף?
כעת קורטיזול משפיע על חילוף החומרים בגוף כאשר הוא נשלט במחזור שינה איטי ויציב שמפחית את ההורמון המשחרר קורטיקוטרופין (CRH) ומגביר את הורמון הגדילה (GH). מחקרי מחקר הראו שכאשר בלוטות יותרת הכליה מפרישות קורטיזול, היא מתחילה לקיים אינטראקציה מורכבת עם ההיפותלמוס ובלוטת יותרת המוח במערכת העצבים והאנדוקרינית. זה גורם לתפקוד יותרת הכליה ובלוטת התריס בגוף להיות קשור קשר הדוק תוך שליטה של ההיפותלמוס וההורמונים הטרופיים. בלוטת התריס מתחרה עם איברי יותרת הכליה על טירוזין. מחקרים מצאו כי טירוזין משמש לייצור קורטיזול במתח תוך מניעת ירידה בתפקוד הקוגניטיבי המגיב ללחץ פיזי. עם זאת, כאשר הגוף אינו יכול לייצר מספיק טירוזין, זה יכול לגרום להיפותירואידיזם ולגרום להורמון הקורטיזול להפוך לכרוני.
סקירה כללית על מתח-וידאו
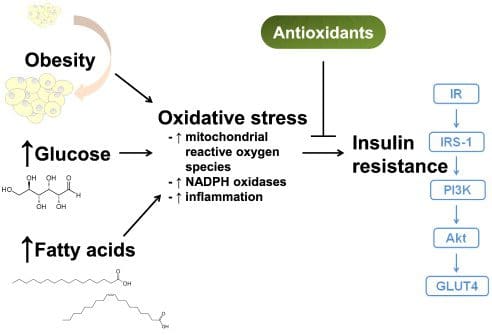
האם חווית כאבי ראש שמופיעים באקראי משום מקום? האם כל הזמן עלית במשקל או ירדת במשקל? האם אתה מרגיש חרד או לחוץ תמיד שזה משפיע על השינה שלך? כל אלה הם סימנים ותסמינים של רמות הקורטיזול שלך שהופכות למצבן הכרוני. הסרטון שלמעלה מראה מה מתח עושה לגוף שלך וכיצד הוא יכול לגרום לתסמינים לא רצויים. כאשר יש סטרס כרוני בגוף, ציר ה-HPA (נוירו-אנדוקריני) אינו מאוזן עקב המפעילים המתווכי-סטרס המעורבים במחלות אוטואימוניות של בלוטת התריס (AITD). כאשר יש לחץ כרוני בגוף, זה יכול לגרום לייצור מוגזם של תרכובות דלקתיות בגוף יכול ליצור IR. החומרים הדלקתיים עלולים לפגוע או להשבית את קולטני האינסולין מה שמוביל לעמידות לאינסולין. זה תורם לפירוק של גורם אחד או יותר הדרושים להשלמת תהליך הובלת הגלוקוז בגוף.
ההשפעות של קורטיזול כרוני בגוף
כאשר יש לחץ כרוני בגוף ולא טופל או מופחת מיד, זה יכול להוביל למשהו המכונה עומס אלוסטטי. עומס אלוסטטי מוגדר כשחיקה של הגוף והמוח עקב פעילות יתר כרונית או חוסר פעילות של מערכות הגוף המעורבות בדרך כלל באתגרים סביבתיים והסתגלות. מחקרי מחקר הראו שעומס אלוסטטי גורם להפרשה עודפת של הורמונים כמו קורטיזול וקטכולמין להגיב לגורמי לחץ כרוניים המשפיעים על הגוף. זה גורם לציר HPA לעשות אחד משני דברים: להיות עמוס יתר על המידה או אי כיבוי לאחר אירועים מלחיצים הגורמים להפרעות שינה. בעיות אחרות שמתח כרוני גורם לגוף יכולות לכלול:
הפרשת אינסולין מוגברת ושקיעת שומן
שינוי בתפקוד החיסוני
תת פעילות בלוטת התריס (תשישות אדרנל)
שימור נתרן ומים
אובדן שנת REM
חוסר יציבות נפשית ורגשית
עלייה בגורמי סיכון קרדיווסקולריים
תסמינים אלו גורמים לגוף להיות לא מתפקד, ו מחקרים הצביעו על כך שגורמי לחץ שונים עלולים לפגוע בגוף. זה יכול להקשות מאוד על אדם להתמודד עם לחץ ולהקל עליו.
סיכום
בסך הכל, מתח או קורטיזול הוא הורמון שהגוף צריך כדי לתפקד כהלכה. מתח כרוני בגוף כתוצאה מגורמי לחץ שונים עלול לגרום להפרעות מטבוליות רבות כמו תת פעילות של בלוטת התריס, עלייה במשקל, תנגודת לאינסולין ותסמונת מטבולית, אם להזכיר כמה. מתח כרוני יכול גם לגרום להפרעות שינה מכיוון שציר ה-HPA מחווט ונראה כאילו הוא נרגע במעט. כאשר אנשים מתחילים למצוא דרכים להתמודד עם גורמי הלחץ השונים הללו, הם יכולים להפחית את רמות הלחץ שלהם בחזרה לנורמליות ולהיות נטולי מתח.
הפניות
ג'ונס, קרול וכריסטופר גוונין. "חוסר ויסות ברמת קורטיזול ושכיחותו - האם זה השעון המעורר של הטבע?" דוחות פיזיולוגיים, John Wiley and Sons Inc., ינואר 2021, www.ncbi.nlm.nih.gov/pmc/articles/PMC7749606/.
McEwen, Bruce S. "השפעות מרכזיות של הורמוני סטרס בבריאות ובמחלות: הבנת ההשפעות המגנות והמזיקות של מתווכי מתח ולחץ." כתב העת האירופי של פרמקולוגיה, הספרייה הלאומית לרפואה בארה"ב, 7 באפריל 2008, www.ncbi.nlm.nih.gov/pmc/articles/PMC2474765/.
Rodriquez, Erik J, et al. "עומס אלוסטטי: חשיבות, סמנים וקביעת ציון באוכלוסיות מיעוט ופערים." Journal of Urban Health: עלון של האקדמיה לרפואה של ניו יורק, ספרינגר ארה"ב, מרץ 2019, www.ncbi.nlm.nih.gov/pmc/articles/PMC6430278/.
תאו, לורן ועוד. "פיזיולוגיה, קורטיזול - Statpearls - מדף הספרים של NCBI." בתוך: StatPearls [אינטרנט]. אי המטמון (FL), הוצאת StatPearls, 6 בספטמבר 2021, www.ncbi.nlm.nih.gov/books/NBK538239/.
מכיוון שהעולם בתנועה מתמדת, אנשים רבים צריכים לסבול מצבים מלחיצים משפיעים על גופם ובריאותם. הגוף צריך הורמונים כמו קורטיזול להמשיך לתפקד כפי שזה משפיע על מערכת החיסון, העצבים, הלב וכלי הדם והשרירים והשלד, אם להזכיר כמה. תפקיד חיוני נוסף שהגוף זקוק לו הוא גלוקוז, הדורש אנרגיה כדי להיות בתנועה מתמדת. מצבים שגורמים לרמות הקורטיזול ולרמות הגלוקוז לעלות בגוף עלולים להוביל לבעיות כרוניות כמו סוכרת ומתח כרוני. זה גורם לאדם להיות אומלל ולהיות במצב רציני אם לא נשלט עליו מיד. המאמר של היום בוחן כיצד קורטיזול וגלוקוז משפיעים על הגוף ואת הקשר השזור בין מתח וסוכרת. הפנה מטופלים לספקים מוסמכים ומיומנים המתמחים בניהול מתח וטיפולים אנדוקריניים עבור אנשים עם סוכרת. אנו מנחים את המטופלים שלנו על ידי הפניה לספקים הרפואיים הקשורים לנו בהתבסס על בדיקתם כאשר הדבר מתאים. אנו מוצאים שחינוך הוא קריטי לשאילת שאלות בעלות תובנות לספקים שלנו. ד"ר אלכס Jimenez DC מספק מידע זה כשירות חינוכי בלבד. כתב ויתור
האם חווית בעיות שינה בלילה? מה לגבי כאבי ראש תכופים המהווים מטרד לאורך כל היום? או האם שמת לב לירידה מוגזמת במשקל או לעלייה במשקל סביב האמצע שלך? חלק מהתסמינים הללו הם סימנים לכך שרמות הקורטיזול והגלוקוז שלך גבוהות ויכולות להשפיע על הגוף שלך. קורטיזול הוא הורמון המיוצר במערכת האנדוקרינית ויכול להועיל או להזיק לגוף אם לא נבדק באופן קבוע. מחקרים הגדירו קורטיזול כאחד הגלוקוקורטיקואידים הבולטים המופרשים עקב תגובת הביוכימיקלים של הגוף, המאופיין על ידי ציר HPA (היפותלמוס-יותרת המוח-אדרנל) מסייע לאירועים קוגניטיביים. עם זאת, כאשר רמות הקורטיזול הופכות לכרוניות בגוף עקב נסיבות שגורמות לגוף להיות לא מתפקד, זה יכול להשפיע באופן משמעותי על אדם ולגרום לחוסר איזון בציר HPA. חלק מהתסמינים שקורטיזול כרוני מוביל לגוף יכולים לכלול:
חוסר איזון הורמונלי
עמידות לאינסולין
עלייה במשקל
עלייה בשומן ה"בטן" הקרביים
תפוקת קורטיזול מוגברת
בעיות חיסוניות
אלרגיות ואסטמה
מפרקים דלקתיים
התאוששות לקויה מפעילות גופנית
מידע נוסף סיפק שהנוכחות של קורטיזול בגוף יכולה לעזור להגביר את זמינות הגלוקוז בדם למוח. כאשר קורטיזול מספק פונקציונליות של איברים, הגלוקוז בדם מספק אנרגיה לגוף.
כיצד קורטיזול וגלוקוז פועלים בגוף
קורטיזול עוזר לעורר גיוס גלוקוז המונית בכבד, ומאפשר לסינתזת חלבון לחסום לדחוף חומצות אמינו לסוכר עבור הגוף. זה ידוע בתור שחרור חומצות שומן שעבר ביולוגית טרנספורמציה לגלוקוז. כאשר זה קורה, זה עוזר לעורר אחסון שומן קרביים אם עודף גלוקוז אינו מנוצל, ובכך גורם לעלייה במשקל. מחקרי מחקר הראו שחוסר בקורטיזול עלול לגרום לירידה בייצור הגלוקוז הכבד בגוף. זה יגרום להיפוגליקמיה, שבה לגוף אין מספיק גלוקוז במערכת שלו. מחקר נוסף מראה שקורטיזול מגיב לכל גורם סטרס שמשפיע על אדם עם רמות גלוקוז נמוכות אך יכול להיות חיובי גם לאחר עומס גלוקוז. ניהול רמות הגלוקוז והקורטיזול בגוף יכול לעזור להתקדם בהתפתחות הסוכרת.
איך קורטיזול קשור לסוכרת סוג 2- וידאו
האם חווית מצבי לחץ שגורמים לשרירים שלך להימתח? מה דעתך להרגיש שרמת הסוכר שלך עולה או יורדת? האם אתה מרגיש תופעות דלקתיות בכל הגוף שגורם להם לכאוב? מתח יכול לגרום להשפעות מזיקות לגוף, להפעיל דלקת, להגביר את הטונוס הסימפטי ולהפחית את ההיענות לגלוקוקורטיקואידים. מתח יכול להיות קשור גם לסוכרת, שכן הסרטון למעלה מראה כיצד הורמון הסטרס קורטיזול קשור לסוכרת מסוג 2. מחקרים הזכירו שקורטיזול יכול להפוך לקשר שלילי למכניקה של תנגודת לאינסולין, להגביר את תפקוד תאי הבטא ולהגדיל את האינסולין המשתחרר בגוף. זה יכול להיות מסוכן עבור אנשים רבים שיש להם סוכרת קיימת והתמודדו עם מתח ללא הרף.
הקשר השזור בין מתח וסוכרת
הקשר השזור בין מתח וסוכרת מוצג כ מחקרים מצאו שהפתופיזיולוגיה של חרדה וסוכרת העלתה את הסיכון לתנגודת אינסולין עבור הגוף. כאשר אדם מתמודד עם מתח כרוני, זה יכול לגרום לו לסבול מבעיות רבות כמו:
חוסר סובלנות קר
קוגניציה ומצב רוח מופחתים
רגישות למזון
אנרגיה נמוכה לאורך כל היום
כאשר זה קורה, הגוף נמצא בסיכון גבוה לפתח עמידות לאינסולין וסוכרת מסוג 2. מחקרים הזכירו שסוכרת מסוג 2 מאופיינת בתנגודת לאינסולין ובתפקוד לקוי של תאי בטא. הגלוקוקורטיקואיד בגוף יכול להיות מוגזם כדי להשפיע על התאים, ולגרום לחוסר תפקוד. מחקרים נוספים הראו שכל מתח נתפס יכול להפוך לגורם סיכון חיוני שלא רק משפיע על הגוף, כמו יתר לחץ דם, BMI (אינדקס מסת גוף), או איכות הדיאטה, אלא יכול לגרום לעלייה בסוכרת מסוג 2. כאשר אנשים מוצאים דרכים להוריד את הלחץ הכרוני שלהם, זה יכול לעזור לנהל את רמות הגלוקוז שלהם מלהגיע לרמות קריטיות.
סיכום
הלחץ הכרוני של הגוף עלול לגרום לתנגודת לאינסולין ולגרום לסוכרת להיות קיימת מראש. הגוף זקוק לקורטיזול וגלוקוז כדי להמשיך לתפקד ולהיות בעל אנרגיה לזוז. כאשר אנשים מתחילים לסבול ממתח כרוני ומסוכרת, זה יכול להיות מאתגר לניהול; עם זאת, ביצוע שינויים קלים בגוף כמו מציאת דרכים להורדת מתח, אכילת מזון בריא וניטור רמות הגלוקוז יכולים לעזור לגוף לאפס את רמות הגלוקוז והקורטיזול למצב נורמלי. פעולה זו יכולה להקל על אנשים רבים שרוצים להמשיך במסע הבריאות שלהם ללא מתח.
הפניות
אדם, Tanja C, et al. "קורטיזול קשור באופן שלילי לרגישות לאינסולין בקרב נוער לטינו עם עודף משקל." כתב העת של אנדוקרינולוגיה קלינית ומטבוליזם, The Endocrine Society, אוקטובר 2010, www.ncbi.nlm.nih.gov/pmc/articles/PMC3050109/.
De Feo, P, et al. "תרומה של קורטיזול לוויסות נגד של גלוקוז בבני אדם." כתב העת האמריקאי לפיזיולוגיה, הספרייה הלאומית לרפואה בארה"ב, יולי 1989, pubmed.ncbi.nlm.nih.gov/2665516/.
Hucklebridge, FH, et al. "תגובת הקורטיזול המתעוררת ורמות הגלוקוז בדם." מדעי חיים, הספרייה הלאומית לרפואה בארה"ב, 1999, pubmed.ncbi.nlm.nih.gov/10201642/.
יוסף, יהושע ג', ושריתה ה' גולדן. "חוסר ויסות קורטיזול: הקשר הדו-כיווני בין מתח, דיכאון וסוכרת מסוג 2." תולדות האקדמיה למדעים של ניו יורק, הספרייה הלאומית לרפואה בארה"ב, מרץ 2017, www.ncbi.nlm.nih.gov/pmc/articles/PMC5334212/.
קמבה, איה ועוד. "קשר בין רמות קורטיזול גבוהות יותר בסרום וירידה בהפרשת אינסולין באוכלוסייה כללית." פלוס אחד, ספרייה ציבורית למדע, 18 בנובמבר 2016, www.ncbi.nlm.nih.gov/pmc/articles/PMC5115704/.
Lee, Do Yup, וכו'. "היבטים טכניים וקליניים של קורטיזול כסמן ביוכימי של מתח כרוני." דוחות BMB, החברה הקוריאנית לביוכימיה וביולוגיה מולקולרית, אפריל 2015, www.ncbi.nlm.nih.gov/pmc/articles/PMC4436856/.
תאו, לורן ועוד. "פיזיולוגיה, קורטיזול." בתוך: StatPearls [אינטרנט]. אי המטמון (FL), הוצאת StatPearls, 6 בספטמבר 2021, www.ncbi.nlm.nih.gov/books/NBK538239.
הגוף הוא מכונה שעובדת היטב שיכולה לסבול כל דבר שנזרק בדרכה. עם זאת, כאשר הוא מקבל פציעה, תהליך הריפוי הטבעי של הגוף יבטיח שהגוף יוכל לחזור לפעילות היומיומית שלו. תהליך הריפוי של שריר פצוע משתנה בכל הגוף. תלוי עד כמה חמור הנזק וכמה זמן ייקח תהליך הריפוי, הגוף יכול להתאושש לכמה ימים עד כמה חודשים בלבד. אחד מתהליכי הריפוי האכזריים ביותר שהגוף צריך לסבול הוא קרע בגיד כף הרגל.
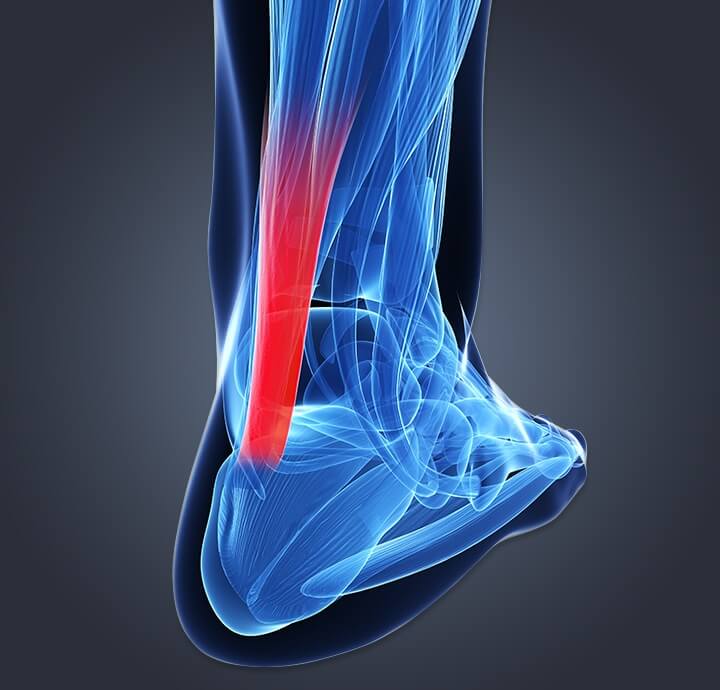
גיד הקלקנאל
גיד השוק או גיד אכילס הוא גיד עבה שנמצא בחלק האחורי של הרגל. גיד השריר הזה הוא מה שגורם לגוף לנוע בזמן הליכה, ריצה או אפילו קפיצה. לא רק זה, גיד השוק הוא הגיד החזק ביותר בגוף, והוא מחבר בין שרירי הגסטרוקנמיוס והסולאוס בעצם העקב. כאשר גיד השוק נקרע, תהליך הריפוי יכול להימשך בין שבועות לחודשים עד להחלמה מלאה.
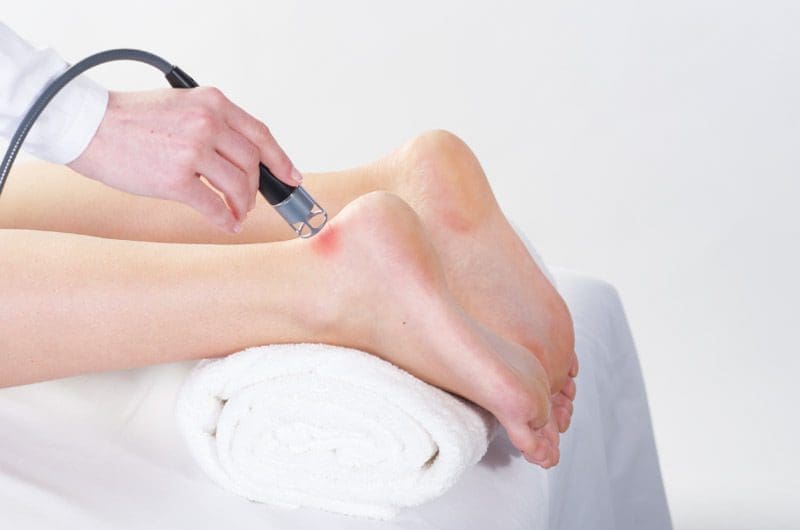
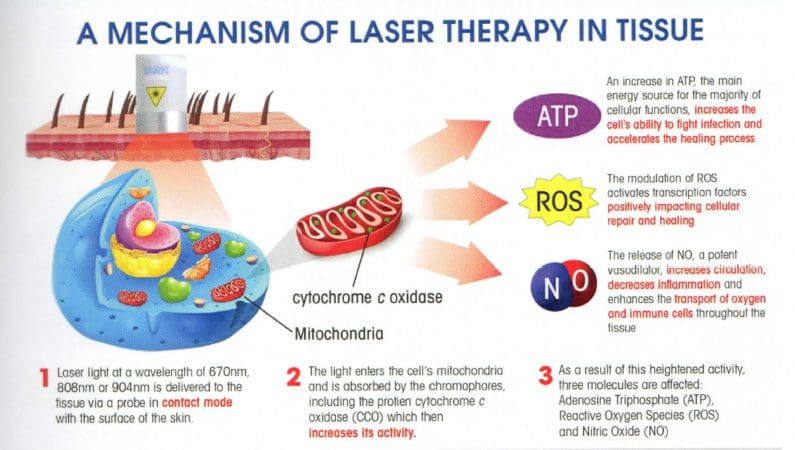
ההשפעות המרפאות של טיפול בלייזר נמוך
אחת הדרכים שיכולות לסייע לתהליך ההחלמה של גידים calcaneal פגומים היא טיפול בלייזר נמוך. מחקרים הראו שטיפול בלייזר נמוך יכול לזרז את תיקון הגיד הפגוע לאחר נגע חלקי. לא רק זה אלא המסרקטיפול באולטרסאונד וטיפול בלייזר נמוך נחקר כגורמים הפיזיים לטיפול בפציעות גידים. המחקרים הראו שלשילוב של טיפול בלייזר נמוך ואולטרסאונד יש תכונות מועילות בתהליך ההחלמה של טיפול בפציעות בגיד השוק.
המחקר מצא שכאשר חולים מטופלים בגידי השוק שלהם, רמות ההידרוקסיפרולין שלהם סביב האזור המטופל מוגברות משמעותית עם אולטרסאונד ו-t laser lowherapy. המבנים הביוכימיים והביומכניים הטבעיים של הגוף על הגיד הפגוע מתגברים, ובכך משפיעים על תהליך הריפוי. מחקר אחר הראה שטיפול בלייזר נמוך יכול לסייע בהפחתת פיברוזיס ולמנוע מתח חמצוני בגיד השוק הפגוע. המחקר אף הראה כי לאחר טראומה של גיד השוק, נוצרות באזור הפגוע דלקת, אנגיוגנזה, הרחבת כלי הדם והמטריצה החוץ-תאית. אז כאשר חולים מטופלים בטיפול לייזר נמוך במשך כארבעה עשר עד עשרים ואחד ימים, החריגות ההיסטולוגיות שלהם מוקלות, ומפחיתות את ריכוז הקולגן והפיברוזיס; מניעת התגברות מתח חמצוני בגוף.
סיכום
בסך הכל, נאמר שההשפעות של טיפול בלייזר נמוך יכולות לעזור להאיץ את תהליך הריפוי של תיקון גיד השוק. התוצאות המבטיחות הוכחו מאחר שטיפול בלייזר נמוך יכול לסייע בתיקון הגיד הפגוע, הפחתת מתח חמצוני ומניעת הסלמה של פיברוזיס, וגורם לבעיות נוספות בגיד הפגוע. ועם השילוב של אולטרסאונד, גיד השוק יכול להתאושש מהר יותר כך שהגוף יוכל להמשיך בפעילות היומיומית שלו ללא פציעות ממושכות.
הפניות:
דמיר, חוסיין ועוד. "השוואה של ההשפעות של טיפולי לייזר, אולטרסאונד וטיפולי לייזר + אולטרסאונד משולבים בריפוי גידים ניסיוני." לייזרים בכירורגיה ורפואה, הספרייה הלאומית לרפואה בארה"ב, 2004, pubmed.ncbi.nlm.nih.gov/15278933/.
פיליפין, לידיאן איזבל, et al. "טיפול בלייזר ברמה נמוכה (LLLT) מונע מתח חמצוני ומפחית פיברוזיס בגיד אכילס שעבר טראומה של חולדה." לייזרים בכירורגיה ורפואה, הספרייה הלאומית לרפואה בארה"ב, אוקטובר 2005, pubmed.ncbi.nlm.nih.gov/16196040/.
Wood, Viviane T, et al. "שינויים ויישור קולגן הנגרמים על ידי טיפול בלייזר ברמה נמוכה ואולטרסאונד בעצימות נמוכה בגיד הקלקנל." לייזרים בכירורגיה ורפואה, הספרייה הלאומית לרפואה בארה"ב, 2010, pubmed.ncbi.nlm.nih.gov/20662033/.
הכלי Find A Practitioner של IFM הוא רשת ההפניות הגדולה ביותר ברפואה פונקציונלית, שנוצרה במטרה לסייע למטופלים לאתר מטפלים ברפואה פונקציונלית בכל מקום בעולם. מטפלים מוסמכים של IFM מפורטים ראשונים בתוצאות החיפוש, לאור השכלתם הרחבה ברפואה פונקציונלית